
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 January 2018 | 10(1): 11210–11217

A new subspecies of the Malayan Bamboo Bat (Chiroptera: Vespertilionidae: Tylonycteris malayana eremtaga) from the Andaman Islands, India
Chelmala Srinivasulu 1, Aditya Srinivasulu 2, Bhargavi Srinivasulu 3 & Gareth Jones 4
4 School of Biological Sciences, Life Sciences Building, 24 Tyndall Avenue, Bristol BS8 1TQ, UK
1 chelmalasrinivasulu@gmail.com (corresponding author), 2 juramaia98@gmail.com, 3 bharisrini@gmail.com, 4 gareth.jones@bristol.ac.uk

Abstract: The bamboo bats belonging to the genus Tylonycteris are unique due to their morphology and ecology, and are known from few locations in South Asia. We collected voucher specimens of Tylonycteris malayana from North Andaman Island, which forms the basis of the first specimen-based record of this species from the Andaman & Nicobar Islands, and the second record from India. Our analyses based on morphometrics, craniodental measurements, bacular morphology and molecular phylogeny based on cytochrome c oxidase subunit I gene indicate that the insular population of T. malayana may have diversified in isolation to differ from the mainland forms, and could represent putative new subspecies, described herein. In light of the new findings, we discuss the importance of conducting detailed study on the specimens of the Lesser Bamboo Bat earlier reported from the Andaman & Nicobar Islands to ascertain their taxonomic status.
Keywords: Bamboo Bat, cryptic diversity, molecular phylogeny, North Andaman Island, Tylonycteris pachypus.
The chiropteran fauna of the Andaman Islands has been the subject of study since 1902 (Miller 1902; Hill 1967; Aul et al. 2014). The family Vespertilionidae is represented by several species, including the bamboo bats of the genus Tylonycteris. Hitherto, only the Lesser Bamboo Bat Tylonycteris pachypus (Temminck, 1840) was reported from the Andaman Islands (Dobson 1876; Hill 1967; Aul 2014). While studying the chiropteran diversity of the Andaman Islands, we collected two specimens of bamboo bats of the genus Tylonycteris from North Andaman Island. Tu et al. (2017) while describing a new species, T. tonkinensis, from northeastern Laos and northern Viet Nam, reassigned the forms of Tylonycteris robustula Thomas, 1915 from northeastern India across mainland Southeast Asia to Tylonycteris malayana Chasen, 1940, and restricted T. robustula s. str. to Sumatra. Detailed study of the specimens from the Andaman Islands, supported by phylogenetic analysis, revealed them to be T. malayana, albeit differing from the nominate form, thus prompting the description of a new insular subspecies.
Study Area and Methods
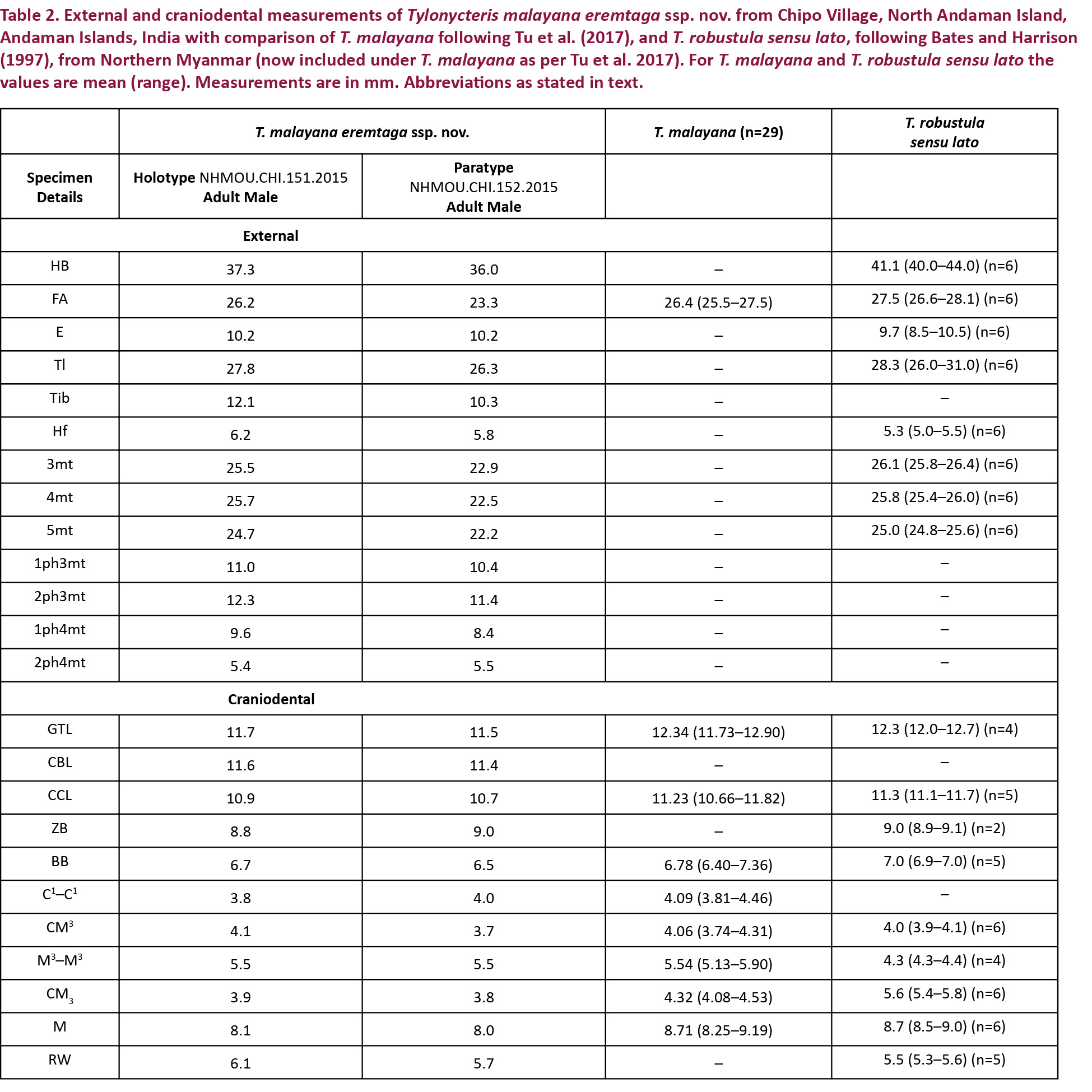
As part of an Indo-UK bat survey and conservation project, we surveyed the Andaman Islands between 2014 and 2016. In November 2015, we mist-netted two small male bamboo bats near Bamboo Tekri, Chipo Village, North Andaman Island, and because we were unsure of their identity, preserved them as vouchers. We took external measurements on live specimens and craniodental measurements of the extracted skulls of the vouchers using digital Vernier calipers (MitutoyoTM, to the nearest 0.01 mm). The following external and craniodental measurements were taken—External: FA, forearm length; E, ear length; Tl, tail length; Tib, tibia length; Hf, hindfoot length; 3mt, third metacarpal; 4mt, fourth metacarpal; 5mt, fifth metacarpal; 1ph3mt, first phalange of third metacarpal; 2ph3mt, second phalange of third metacarpal; 1ph4mt, first phalange of fourth metacarpal; 2ph4mt, second phalange of fourth metacarpal; Craniodental: GTL, greatest length of the skull; CBL, condylobasal length; CCL, condylocanine length; CM3, maxillary toothrow; C1–C1, anterior palatal width; M3–M3, posterior palatal width; ZB, zygomatic breadth; BB, braincase breadth; CM3, mandibular toothrow; M, mandible length. Bacula were extracted and stained following the standard method (Topal 1958). We compared our specimens with two specimens of T. malayana (labelled as T. robustula pre-Tu et al. 2017) from Malaysia, housed in the Harrison Zoological Museum, UK (specimen nos. 2.7443 and 3.7444), and also with external and craniodental measurements of the nominate form provided by Vuong Tan Tu (pers. comm., October 31, 2017).
Wing punches from the specimens were taken and preserved in silica gel. Genomic DNA was then extracted using DNEasy Blood and Tissue kit (QIAGEN). A PCR was conducted to amplify partial cytochrome c oxidase subunit 1 (COI) gene sequences using forward and reverse primers VF1d (5’- TTCTCAACCAACCACAARGAYATYGG-3’) and VR1d (5’- TAGACTTCTGGGTGGCCRAARAAYCA-3’) (Ivanova et al. 2006). The PCR reaction was performed in a 25μl reaction volume containing 2μl of template DNA, 12.5μl of 2X reaction buffer (0.05U/μL Taq DNA polymerase, reaction buffer, 4mM MgCl2, 0.4mM of each dNTPs), 0.5μl of each primer, and 9.5μl nuclease free water. The thermal profile followed included: 940C for 1 min, five cycles of 940C for 30s, 500C for 40s, and 720C for 1 min, followed by 35 cycles of 940C for 30s, 550C for 40s, and 720C for 1 min, with a final extension at 720C for 10 min. The amplified PCR products were sequenced using an ABI prism 3730 sequencer (Applied Biosystems, USA) and big dye terminator sequencing kit (ABI Prism, USA). The gene sequence (633 base pairs, GenBank accession number MG807173) obtained in the study was analyzed using BLAST (Altschul et al. 1990) to double-check the quality of the sequence. Additional sequences of other related species were retrieved from the NCBI GenBank database for phylogenetic analysis (Table 1). The sequences were aligned using MUSCLE (Edgar 2004) incorporated in MEGA6 (Tamura et al. 2013) using default parameters. JModelTest 2 was used to choose the best-fitting maximum likelihood DNA substitution model for the dataset, based on Bayesian Information Criterion (BIC) scores for each model type (Guindon & Gascuel 2003; Darriba et al. 2012). The analysis used the Hasegawa-Kishino-Yano + Gamma [ncat=4] (HKY+G, BIC = 5144.08) nucleotide substitution model (Hasegawa et al. 1985). We constructed a Bayesian Inference of phylogeny in the BEAST 1.8.2 package (Drummond et al. 2012), with default priors, using the nucleotide substitution model obtained for a chain length of 10 million generations, sampling every 1000 generations. The tree was rooted using Pipistrellus javanicus and Hypsugo cadornae as outgroup taxa. We double-checked convergence by plotting likelihood scores against generations in Tracer v1.6 (Rambaut et al. 2014). The first 20% of the trees were discarded as burn-in, and chronograms were created using TreeAnnotator v1.8.2 (as part of the BEAST package) and visualized in FigTree v1.4.1 (Rambaut 2009). Kimura 2-parameter (K2P) pairwise distances were calculated using MEGA6.
Results
We confirmed the identity of the specimens as T. malayana, based on external and craniodental measurements, diagnostic characters, and molecular phylogeny using partial cytochrome c oxidase subunit I gene sequence. Owing to discernible variation in morphological and craniodental measurements, bacular structure, and genetic distance, we describe a new insular subspecies of T. malayana.
Tylonycteris malayana eremtaga ssp. nov.
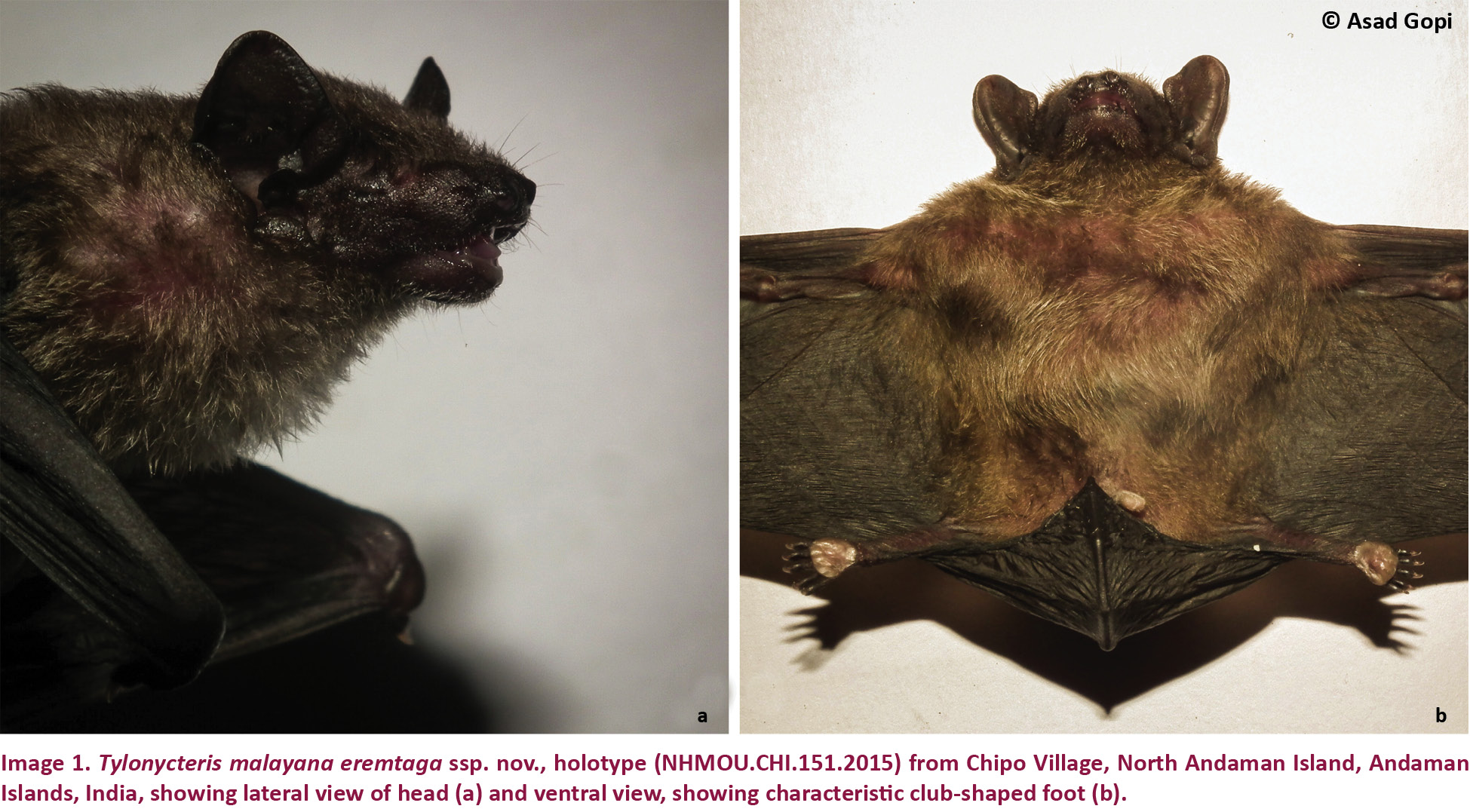
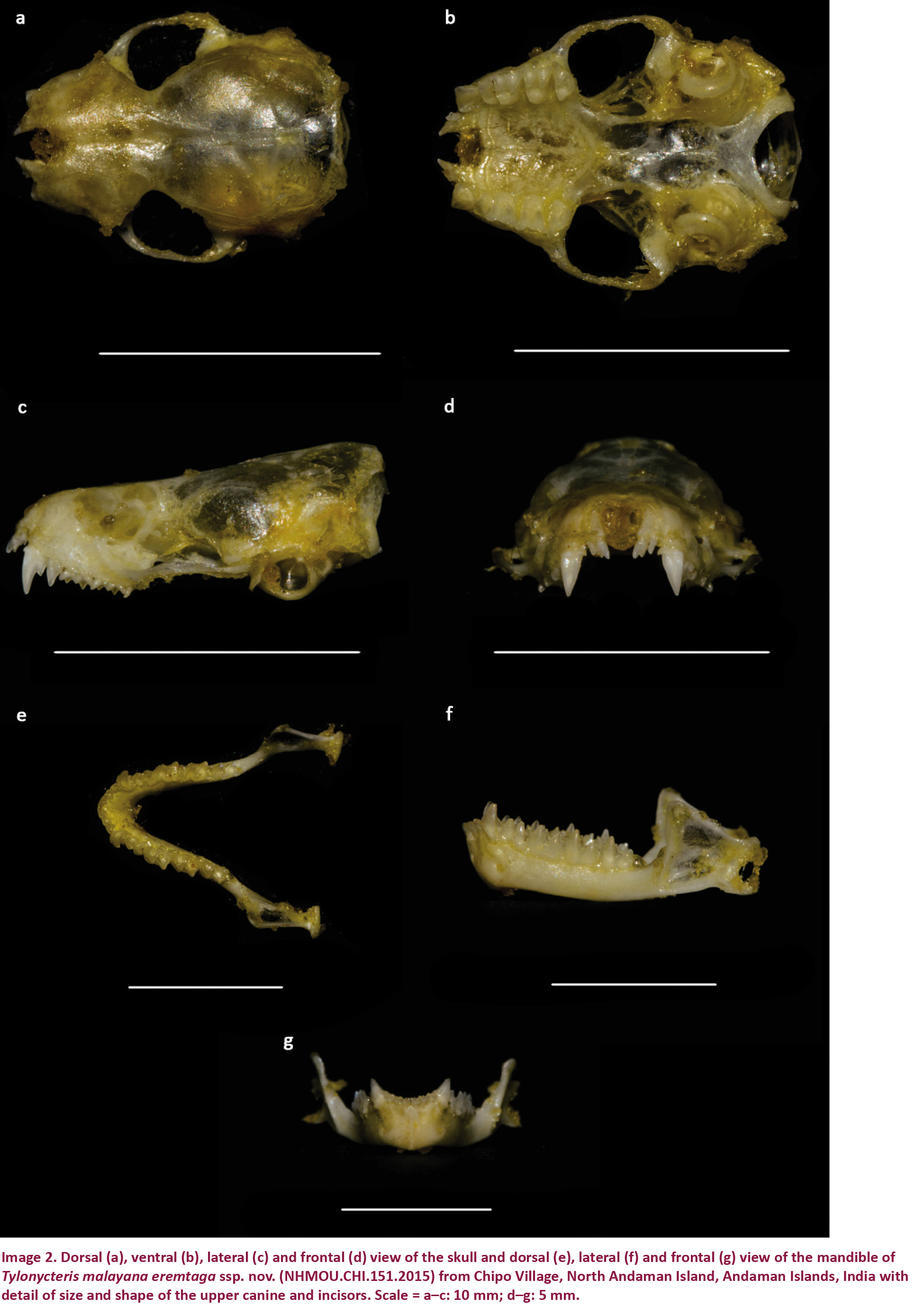
(Images 1a,b, 2a–g, 3a–c)
urn:lsid:zoobank.org:act:003A365F-B5E5-4EE8-B150-C7E6D8000731
Andaman Bamboo Bat
Holotype: NHMOU.CHI.151.2015, adult male, 06.xi.2015, near Bamboo Tekri (13.3730N & 92.9990E), Chipo Village, Diglipur, North Andaman Island, Andaman & Nicobar Islands, India; coll. A. Gopi & Tauseef Hamid Dar.
Paratype: NHMOU.CHI.152.2015, adult male, other details same as holotype.
Diagnosis: A small bat with a forearm length up to 23.3–26.25 mm. Dorsal pelage uniformly grey-brown, venter slightly paler; wings and interfemoral membrane dark brown. Although the new subspecies is essentially similar to nominotypic T. m. malayana (see Tu et al. 2017), it varies from the nominate form slightly in having smaller craniodental features (GTL: 11.5 in T. m. eremtaga ssp. nov. vs 11.73–12.90 in T. m. malayana). The rostrum is robust in T. m. eremtaga ssp. nov., shorter and broader than that of the nominate subspecies (CM3: 3.7–4.1 in T. m. eremtaga ssp. nov. vs 3.74–4.31 in T. m. malayana; RW: 5.7–6.1 in T. m. eremtaga ssp. nov. vs 5.3–5.6 in T. robustula sensu lato (see Bates & Harrison 1997). Externally, the fur color of T. m. eremtaga ssp. nov. appears to be uniform grey-brown while that of T. m. malayana is dark brown.
Description: A small bat with a forearm length up to 26.25mm. Head characteristically flat, projecting forwards and downwards (Image 1a). Ears roughly triangular with broadly rounded tips. Tragus short and broad. Base of the thumb and the soles of the feet have broad fleshy pads (Image 1b). Wings short with 3rd, 4th and 5th metacarpals roughly equal in length. Wing and interfemoral membranes dark brown (Image 1a,b), pelage fine and dense, uniformly grey-brown on the dorsum, slightly paler on the ventral surface.
The skull is small, with a length (GTL) up to 11.7mm. The whole skull is characteristically flattened and broad (Image 2a,c,d). The breadth of the braincase exceeds twice the occipital height, and the rostrum is short and broad (Image 2a), with distinct supraorbital projections. Sagittal crest absent. Lambdoid crest weakly developed. Mesopterygoid space wide and short. Horizontal ramus of each half-mandible is relatively thick, coronoid process large, condyle and angular process well developed (Image 2f).
Upper toothrow length (CM3) up to 3.8mm. The first incisor (i3) is large, narrow and bicuspidate; second is unicuspidate and roughly equal to the length of the outer cusp of i2. The upper canine is separated from i3 by a short diastema, and is double the length of i2. Upper canine has a distinct posterior secondary cusp. Upper premolar (pm4) has a crown area approximately half that of m1 and slightly exceeds half of the height of the upper canine. M1 slightly exceeds m2 in breadth, and has a distinct protocone. M3 lacks a metastyle but it is not greatly reduced. The three lower incisors are tricuspidate and located adjoining the lower canine. Lower canine weak, not as robust as the upper canine and slightly exceeding the protoconid of m2 in height. Pm2 and pm4 are equal in size and compressed in the toothrow, anterior part of pm2 positioned on the posterior cingular cusp of the canine. In m1 and m2, the talonid exceeds the trigonid in size, in m3 they are roughly equal.
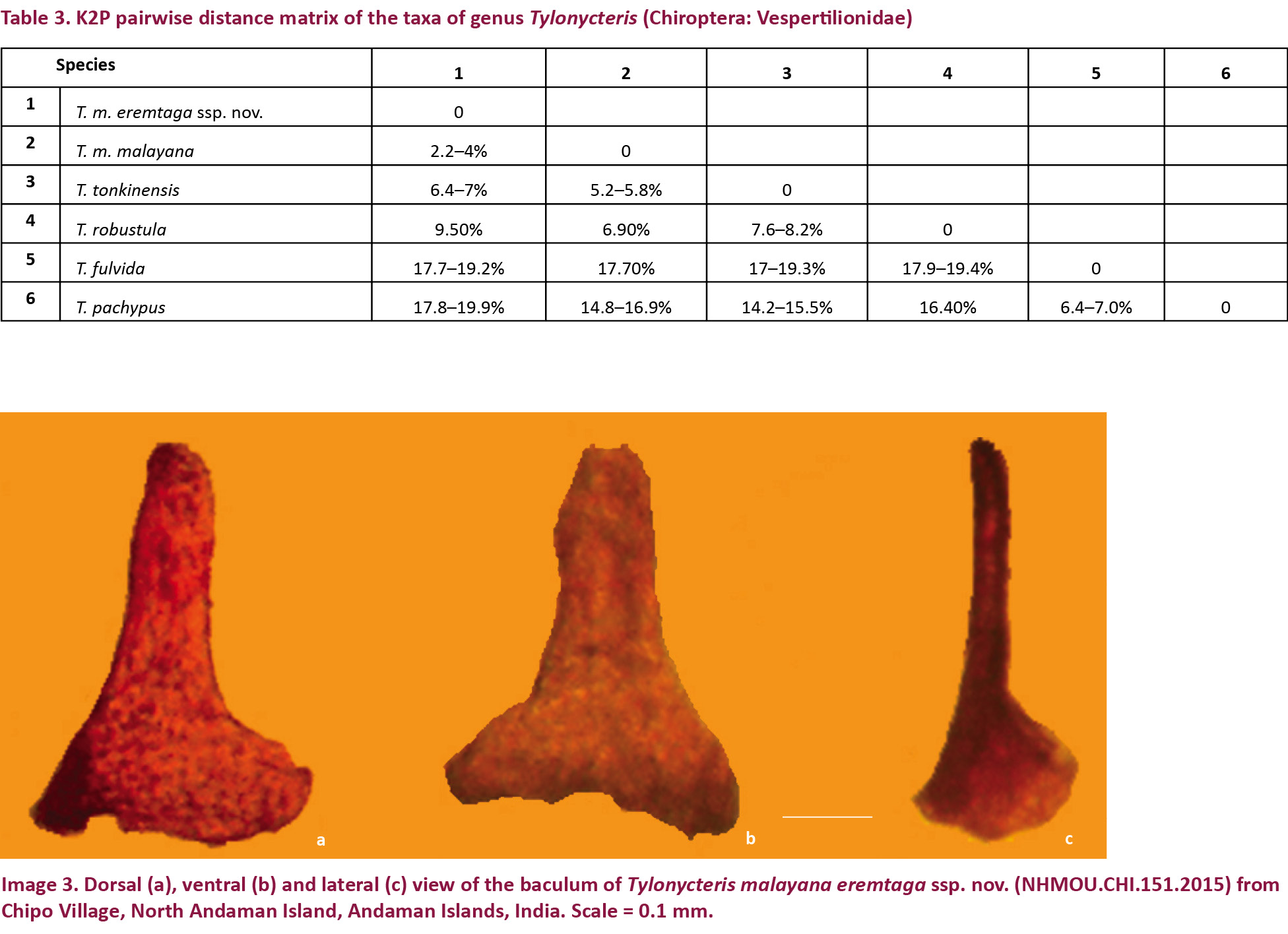
The baculum of the new subspecies is located at the tip of the short penis, which has a globular glans. It has a broad, flared base (0.3mm) (Image 3), which is concave in the holotype and slightly notched in the paratype. The shaft of the baculum of the holotype is long (0.4mm in length), straight, with a slight ventral concavity beneath the broadly rounded tip. In the lateral aspect, the shaft of the baculum is long and straight, and the base is flared, unlike that of the nominate subspecies, in which the shaft curves dorsally along its length, and the base is knob-like (Hill & Harrison 1987). The paratype matches the holotype in all characters excepting size (Table 2).
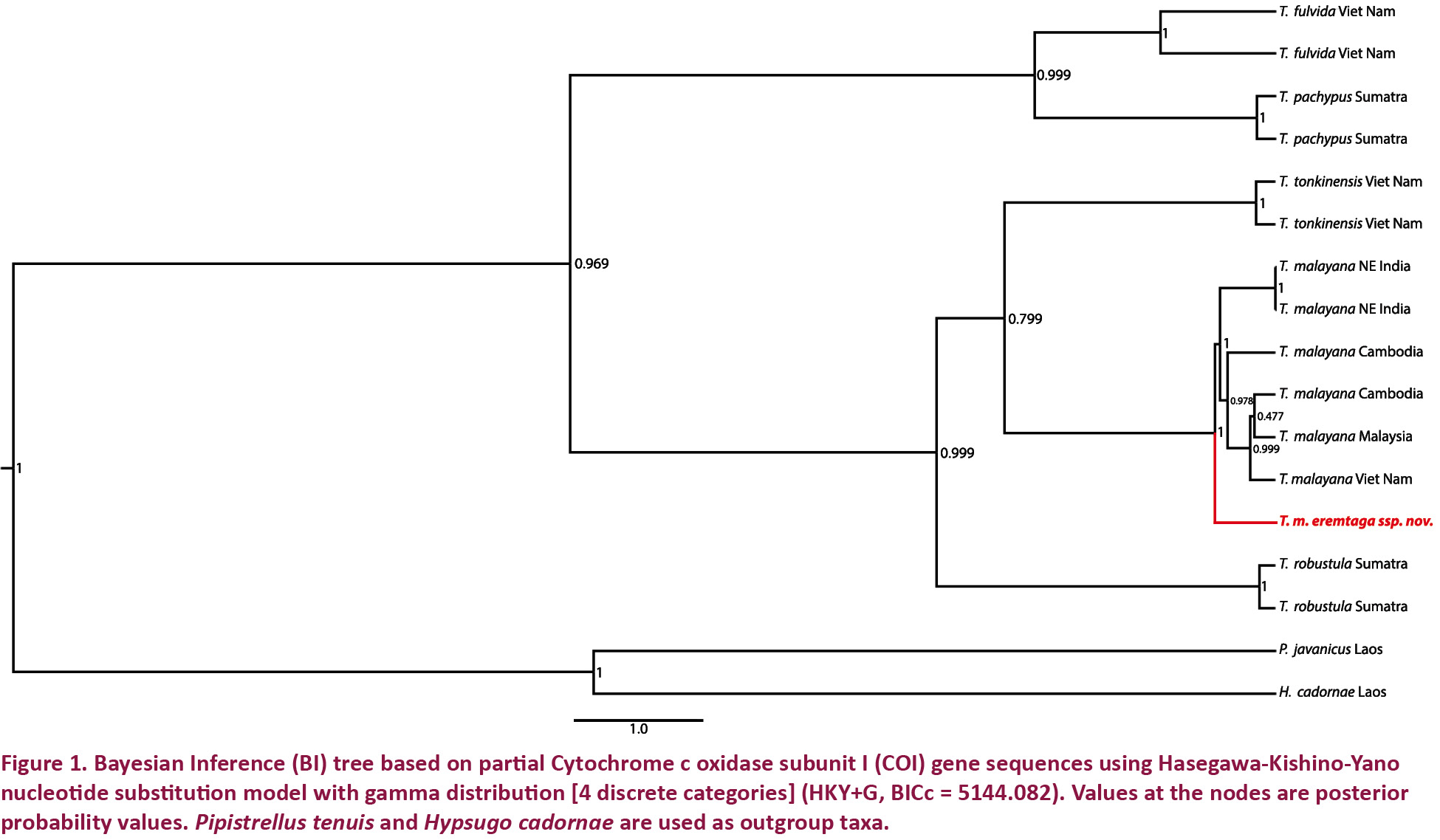
Molecular phylogeny: The phylogenetic tree obtained from the analysis of COI gene of the Tylonycteris species supports the distinctness of the Andaman population with the taxa previously assigned to T. robustula sensu lato and T. pachypus sensu lato clading separately with high posterior support, in agreement with Tu et al. (2017). Within the T. robustula sensu lato clade, five distinct subclades were observed: one subclade of T. robustula sensu stricto from Sumatra, one subclade of T. tonkinensis from Laos and Viet Nam, two separate subclades of T. m. malayana from Southeast Asia and India respectively, and one subclade of T. m. eremtaga ssp. nov. from the Andaman Islands, supported by high posterior probability (> 0.75) (Fig. 1). The distinctness of the new subspecies is also supported by K2P distances of 2.2–4% from the nominate subspecies for the COI gene (Table 3).
Etymology: The subspecific epithet, ‘eremtaga,’ is a Latinized noun derived from the Aka-Kora dialect of the Great Andamanese language, meaning ‘forest-dweller’.
Ecological notes: The presence of multiple males in the area suggests the existence of a colony of these bats in the near vicinity of the village. The specimens collected from Chipo Village, Diglipur, North Andaman Island were found to share their habitat with other species such as the Greater Short-nosed Fruit Bat Cynopterus sphinx, the Lesser Short-nosed Fruit Bat Cynopterus brachyotis, Tickell’s Bat Hesperoptenus tickellii, and the Andaman Intermediate Horseshoe Bat Rhinolophus affinis andamanensis. This subspecies is currently known only from Chipo Village, Diglipur, North Andaman Island, Andaman & Nicobar Islands, India.
Discussion
One taxon, Tylonycteris pachypus sensu lato, has so far been reported to be present on Andaman Islands, from Wimberleygunj, South Andaman Island (Hill 1967), and Webi and Karmatang in North Andaman Island (Aul 2014), principally based on external morphology and forearm length. In light of the recent taxonomic revision (Tu et al. 2017), and the discovery of the T. malayana eremtaga ssp. nov. from the Andaman Islands, it is imperative that the specimens collected earlier from the Andaman Islands (Dobson 1876; Hill 1967; Aul 2014) need to be re-examined further to ascertain their correct identity.
The bamboo bats belonging to genus Tylonycteris depend on bamboo forests, and harvesting of bamboo for commercial purpose, destruction of bamboo forests, forest fire, and increased use of pesticides in fringe villages might have a negative impact on the survival of these species.
References
Altschul, S.F., W. Gish, W. Miller, E.W. Myers & D.J. Lipman (1990). Basic local alignment search tool. Journal of Molecular Biology 215(3): 403–410; http://doi.org/10.1016/S0022-2836(05)80360-2
Aul, B. (2014). Record of Tylonycteris pachypus (Lesser Bamboo Bat) from Andaman Islands. Ambient Science 1(1): 44–46; http://doi.org/10.21276/ambi.2014.01.1.nn03
Aul, B., P.J.J. Bates, D.L. Harrison & G. Marimuthu (2014). Diversity, distribution and status of bats on the Andaman and Nicobar Islands, India. Oryx 48(2): 204–212; http://doi.org/10.1017/S0030605312000646
Bates, P.J.J. & D.L. Harrison (1997). Bats of the Indian Subcontinent. Harrison Zoological Museum, Sevenoaks, Kent, UK. 258pp.
Darriba, D., G.L. Taboada, R. Doallo & D. Posada (2012). jModelTest 2: more models, new heuristics and parallel computing. Nature Methods 9(8): 772; http://doi.org/10.1038/nmeth.2109
Dobson, G.E. (1876). Monograph of the Asiatic Chiroptera and catalogue of the species of bats in the collection of the Indian Museum, Calcutta. Indian Museum, London.
Drummond, A.J., M.A. Suchard, D. Xie & A. Rambaut (2012). Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Molecular Biology and Evolution 29(8): 1969–1973; http://doi.org/10.1093/molbev/mss075
Edgar, R.C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research 32(5): 1792–1797; http://doi.org/10.1093/nar/gkh340
Guindon, S. & O. Gascuel (2003). A simple, fast and accurate method to estimate large phylogenies by maximum-likelihood. Systematic Biology 52: 696–704; http://doi.org/10.1080/10635150390235520
Hasegawa, M., H. Kishino & T. Yano (1985). Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. Journal of Molecular Evolution 22(2): 160–174; http://doi.org/10.1007/BF02101694
Hill, J.E. (1967). The bats of Andaman and Nicobar Islands. Journal of the Bombay Natural history Society 64: 1–9.
Hill, J.E. & D.L. Harrison (1987). The baculum in the Vespertilioninae (Chiroptera: Vespertilionidae), with a systematic review, a synopsis of Pipistrellus and Eptesicus, and the descriptions of a new genus and subgenus. Bulletin of the British Museum (Natural History) Zoology 52: 225–305.
Ivanova, N.V., J.R. Dewaard & P.D.N. Hebert (2006). An inexpensive, automation-friendly protocol for recovering high-quality DNA. Molecular Ecology Notes 6: 998–1002; http://doi.org/10.1111/j.1471-8286.2006.01428.x
Miller, G.S. (1902). The mammals of the Andaman and Nicobar Islands. Proceedings of the United States National Museum 24: 751–795.
Rambaut, A. (2009). FigTree version 1.4 [computer program] http://tree.bio.ed.ac.uk/
Rambaut, A., M.A. Suchard, D. Xie & A.J. Drummond (2014). Tracer version 1.6 [computer program] http://tree.bio.ed.ac.uk/software/tracer/
Tamura, K., G. Stecher, D. Peterson, A. Filipski & S. Kumar (2013). MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Molecular Biology and Evolution 30(12): 2725–2729; http://doi.org/10.1093/molbev/mst197
Topal, G. (1958). Morphological studies on the os penis of bats in the Capathian Basin. Annales historico-naturales Musei nationalis hungarici 50(New series 9): 331–342.
Tu, V.T., G. Csorba, M. Ruedi, N.M. Furey, N.T. Son, V.D. Thong, C. Bonillo & A. Hassanin (2017). Comparative phylogeography of bamboo bats of the genus Tylonycteris (Chiroptera, Vespertilionidae) in Southeast Asia. European Journal of Taxonomy 274: 1–38; http://doi.org/10.5852/ejt.2017.274
