Journal of Threatened Taxa | www.threatenedtaxa.org | 26 January 2018 | 10(1): 11147–11155

Population characteristics of Silaum silaus (L.) Schinz & Thell. (Apiaceae) in Mordovia, a highly threatened plant species at the northern limit of its range
Anatoliy A. Khapugin
Joint Directorate of the Mordovia State Nature Reserve and National Park “Smolny”, 430011, Saransk,
Republic of Mordovia, Russia
hapugin88@yandex.ru

Abstract: Knowledge on status of peripheral plant populations is important for understanding of species’ adaptation and evolution within their ranges. There is a lack of data on the status of Silaum silaus (Apiaceae) populations at the periphery of its native range. One of the most northern native S. silaus populations is situated in the Republic of Mordovia (Central Russia). This species is considered as Critically Endangered in the region. Population-based studies of S. silaus have been carried out. Reproductive biology, morphometric parameters of individuals, accompanying flora, seed characteristics (mass, germination rate) were investigated. Results showed that at the northern limit of the range S. silaus grows in floodplain meadow community previously disturbed by livestock grazing. The area occupied by the population, however, has increased more than 12 fold over the last 15 years. Increasing fruit production together with decreasing fruit mass was established for the studied population compared to populations in the central part of its native range. In addition, germination rate was very low (2.3–16.7 % depending on edaphic conditions). Additional studies of S. silaus populations are needed in saline steppes of the closely located territories where it occurs as one of the dominant species in the plant community.
Keywords: Accompanying flora, Apiaceae, endangerment, habitat conditions, peripheral population, phytoindication, reproductive biology, seed ecology, seed germination.
Russian abstract: Знания о состоянии периферических популяций растений очень важны для понимания адаптации и эволюции видов в пределах их ареалов. В настоящее время имеется недостаток данных о состоянии популяций Silaum silaus (Apiaceae) на границе естественного его распространения. Одна из самых северных аборигенных популяций S. silaus расположена в Республике Мордовия (Центральная Россия). S. silaus рассматривается в этом регионе как таксон, находящийся на грани исчезновения. Были проведены популяционные исследования S. silaus. Были изучены репродуктивная биология, морфометрические параметры особей, состав сопутствующей флоры, характеристики семян (вес, всхожесть). Полученные результаты показали, что на северной границе ареала S. silaus произрастает в пойменно-луговом сообществе, пострадавшем от выпаса скота. Несмотря на это, площадь популяции за последние 15 лет увеличилась более, чем в 12 раз. Для изученной популяции было установлено увеличение продуктивности семян совместно с уменьшением их массы по сравнению со значениями этих параметров в центральной части естественного ареала этого вида. Кроме того, всхожесть семян S. silaus была очень низка (от 2.3% до 16.7% в зависимости от эдафический условий).Необходимы дополнительные исследования популяций S. silaus в области засоленных степных сообществ в соседних регионах, где этот вид наблюдается в качестве одного из доминантов.
Introduction
The genus Silaum Mill. (Apiaceae) consists of one species Silaum silaus (L.) Schinz & Thell. (Pimenov & Tikhomirov 1979; The Plant List 2013) distributed from the Iberian peninsula in the southwest to western Siberia in the east. The north range’s boundary passes through southern Sweden (Tutin 1968; Menglan & Watson 2005; Pimenov & Ostroumova 2012). In some regions of Europe S. silaus was introduced with material coming from Germany during World War II (Fröberg 2009). The species was noted as alien in Jiangsu Province, on the eastern coast of China (Menglan & Watson 2005).
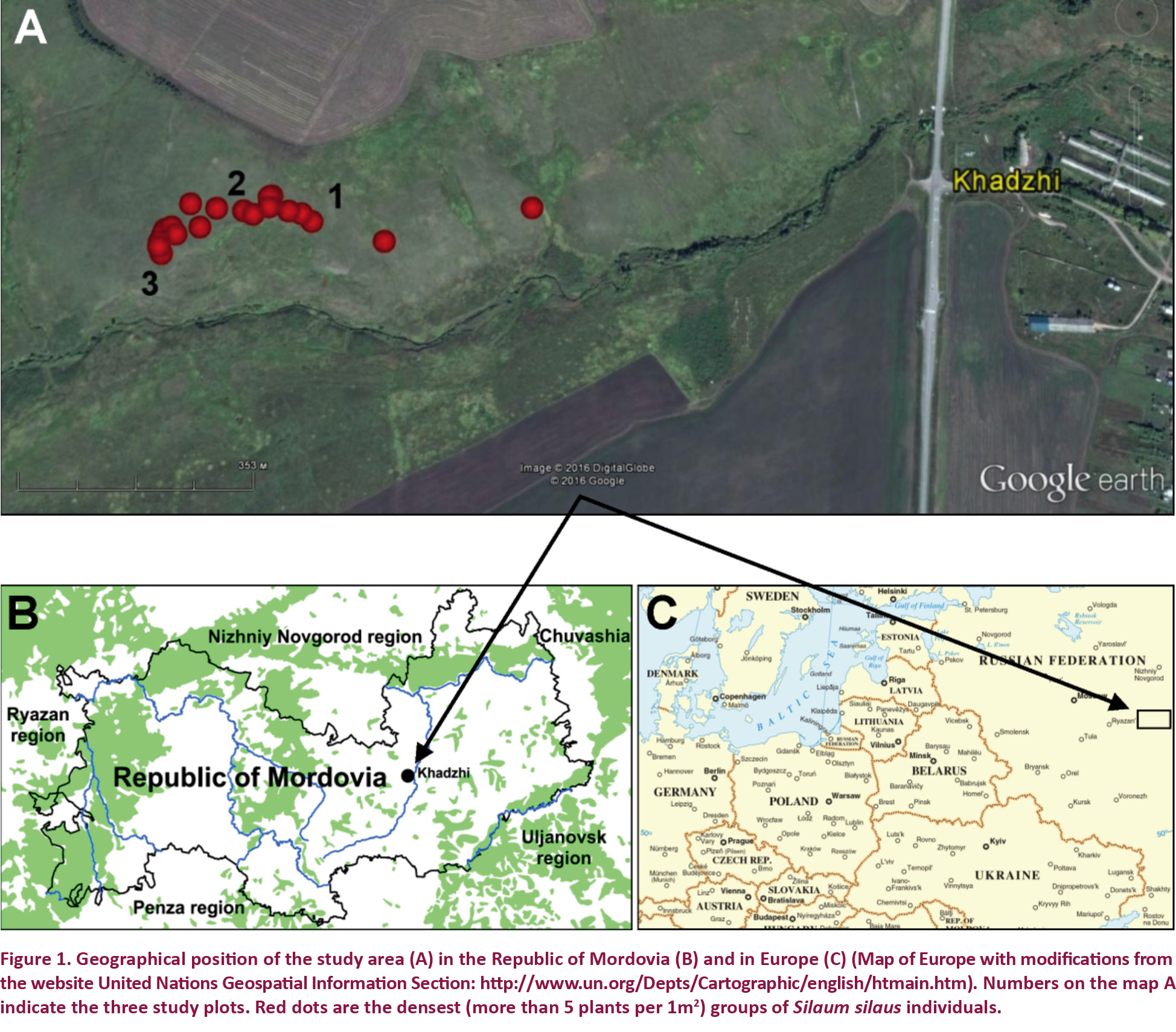
In Russia native S. silaus populations are known in 26 regions in the middle part of European Russia. These include: Belgorod, Voronezh, Lipetsk, Penza, Tambov, Samara, Saratov, Uljanovsk, Chuvashia and Mordovia. This species is considered as alien in: Irkutsk, Leningrad and Tver (Pimenov & Ostroumova 2012; Tikhomirov 2014). Finally, one of the most northern native S. silaus populations in Central Russia is situated near Khadzhi Village in Lyambir District of the Republic of Mordovia (Fig. 1).
Silaum silaus is a perennial Apiaceae with annual shoots up to 40–100 cm tall (Tutin 1968; Menglan & Watson 2005; Pimenov & Ostroumova 2012) (Image 1). Its main characteristics are as follows: leaf blade triangular-ovate, 7–20 × 6–10 cm; lateral pinnae short-petiolulate, 2–5-lobed, terminal pinnae 3–7-lobed; ultimate segments lanceolate to linear-lanceolate, 13–20 × 2–3 mm, abaxial veins prominent, margins cartilaginous, apex acute or acuminate, apiculate. Upper leaves bipinnate, narrowly linear, apical leaves reduced, segments filiform. Umbels 2.5–4.0 cm across; rays 5–10, 1–3 cm, unequal; bracteoles linear-lanceolate, 3–5 mm, shorter than flowers, margin scarious; umbellules many-flowered; pedicels 4–8 mm. Fruits 4–8 × 2–3 mm (Image 1). Within its native range it inhabits floodplain grasslands (e.g., Hölzel & Otte 2004; Borovik 2011; Guardiola et al. 2014; Willner et al. 2013). In some cases, S. silaus can dominate plant communities in salinised meadows (e.g., Novikova & Pankina 2013). This species, however, is considered as vulnerable due to limited dispersal and re-establishment even under favourable environmental conditions (Bischoff 2000, 2002; Donath et al. 2006; Bischoff et al. 2009).
In 2002, Silaum silaus was recorded near Khadzhi Village in Lyambir District of the Republic of Mordovia (GMU - Herbarium of the Mordovia State University). At that time dense S. silaus population occupied 800m2. Besides, since 1994 from one to 10 individuals were observed on the railway bed in Ruzaevka City (Silaeva & Barmin 1995). Recently, the regional status of S. silaus was assessed according to the IUCN Red List of Threatened Species categories and criteria (IUCN 2012a,b, 2014) as Critically Endangered (CR) (Khapugin et al. 2017a). Knowledge on structure and status of both peripheral and central plant populations and their conservation status are important to understand the adaptation and evolution within the native range (e.g., Wróblewska & Brzosko 2006; Sotek 2007; Beatty et al. 2008; Abeli et al. 2014; Khapugin & Chugunov 2015; Naik et al. 2016; Srivastava et al. 2017; Birkeland et al. 2017). Such understanding may be achieved through large-scale population-based studies of plants both in the centre and at the limits of their ranges. Until now, however, studies of peripheral S. silaus populations have not been carried out either in Mordovia or in any other regions at the limit of species’ range.
The aim of this paper was to study the peripheral S. silaus population known in the Republic of Mordovia. Two main tasks were established: (i) to reveal current status of S. silaus population; (ii) to compare data obtained with those from populations in the centre of its range (wherever possible).
Materials and Methods
Investigations have been carried out in the grassland community located in the valley of Lyambirka stream at the northern limit of S. silaus’ range (Republic of Mordovia), 54.330N & 45.230E. This location was recommended for the establishment of a protected area (Khapugin et al. 2017b). Current population area has been estimated using portative GPS-navigator. At present, S. silaus population occupies 9,876m2. The population-based study of S. silaus was carried out in August 2016. Three 10×10 m plots were randomly established in eastern, central and western parts of the population (see numbers in Fig. 1A). Red dots in Fig. 1A indicate places with highest plant density.
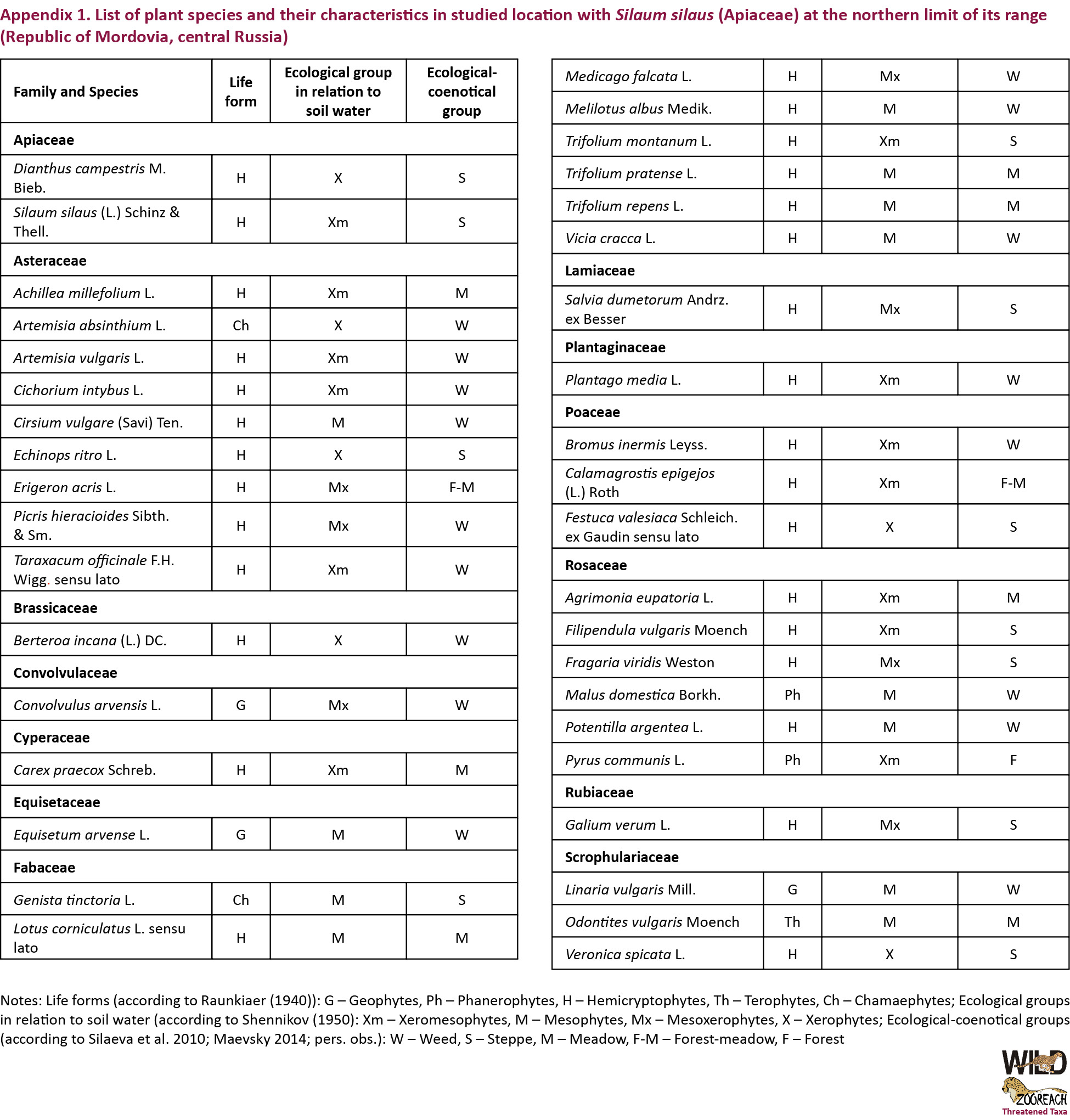
Environmental conditions typical for habitats with S. silaus were estimated using the data on environmental preferences of vascular plant species, accompanying the endangered species in the studied location. For this purpose, all angiosperm species were recorded and these data were analysed within each study plot. Biomorphological analysis of the accompanying flora was done to define species groups according to Raunkiaer’s life-form classification (Raunkiaer 1934). Distribution of plant species of the accompanying flora along the ecological groups in relation to the water content in soil was carried out following the classification of Shennikov (1950) by defining of main mesophytes (plants growing on moderately moisturised soils), xerophytes (plants growing on poorly moisturised soils) and intermediate (xeromesophytes, mesoxerophytes) groups. Ecological-coenotical characteristics for each plant species that was registered in study units were determined using the literature on the flora of the Republic of Mordovia (Silaeva et al. 2010), the flora of Central Russia (Maevsky 2014) and own personal observations.
Concomitantly, values of environmental factors were calculated for the studied habitat based on the accompanying flora composition. Calculations were done according to a scale where environmental indicator values (EIVs) are presented as intervals (Tsyganov’s 1983). It means that we can define the range of each plant existence in relation to certain environmental factors; for example, soil nitrogen, moisture, etc. Mean EIVs were calculated using the algorithm suggested by Buzuk & Sozinov (2009). Ten EIV scales were used: thermo-climatic (TM), climate continentality (KN), climate humidity (OM), kryo-climatic (CR), soil moisture (HD), soil trophicity (TR), soil nitrogen (NT), soil pH (RC), shading (LC), soil moisture variability (FH). These calculated mean EIVs were used to characterise the conditions of the habitat with S. silaus.
Morphometrical parameters of S. silaus individuals (shoot height, number of generative shoots per individual, number of umbels, number of umbellules per umbel, number of fruits per umbellule) were measured. Number of leaves per vegetative individual was determined.
In order to estimate seed characteristics, all S. silaus seeds were collected from 30 randomly sampled individuals (10 umbellules from each plant). They were dried under sunlight. The seed mass was then measured using 20 samples of 50 randomly selected seeds weighed to the nearest 0.1mg. The seeds were then kept in a refrigerator at 3–4 0C for six months (cold stratification). They were sowed in growth chambers in six replications of the experiment, 50 seeds in each of three media (water, sand and soil) under t = 20–25 0C, artificial lighting, 16/8 h (2000 lux).
The seeds were considered as germinated when the radicle was longer than the seed, while the shoot was longer than half of the seed (Nikolaeva et al. 1985). Each experimental replication was conducted until the end of seed germination in the growth chamber. Both the period from seed sowing to their germination and the germination period were recorded. The energy period, germination energy, and seed germination were determined. The energy period was determined experimentally through the comparison of seed germination evenness. For this purpose, the maximum number of germinated seeds per day was recorded. The germination energy was determined as a ratio of the maximum number of germinated seeds per day to the total number of germinated seeds in a replication. Statistical analyses were carried out using PAST 3.15 (Hammer et al. 2001) and Microsoft Excel.

Results
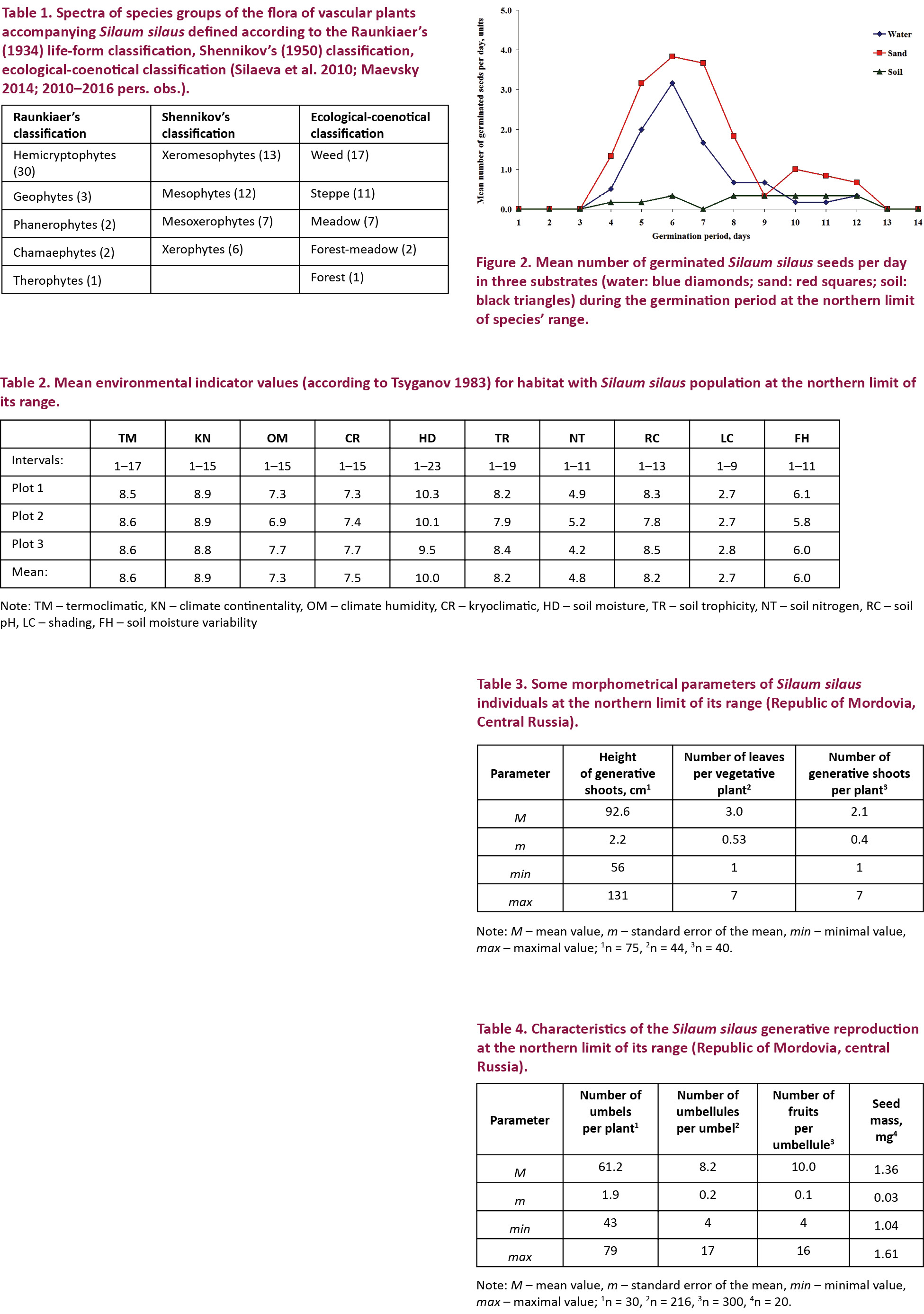
The diversity of vascular plants of this study site comprised 38 vascular plant species belonging to 35 genera from 14 families (Appendix 1). The following species had the highest cover at the different sampled sites: Bromopsis inermis (Leyss.) Holub (up to 80%), Fragaria viridis Weston (up to 60–65 %), Echinops ritro L. (up to 25–30 %), Carex praecox Schreb. (up to 15%), Trifolium pratense L. or even Silaum silaus (both - up to 6%). Amongst species of accompanying flora, hemicryptophytes dominated significantly, as well as submesophytes while subxerophytes prevailed among groups in relation to edaphotope moisture conditions (Table 1). In ecological-coenotical spectra, weed group (including weed and meadow weed subgroups) had the highest species richness (Table 1).
Mean values of environmental factors were calculated according to Tsyganov’s (1983) environmental scales (Table 2). On their basis some environment characteristics of the habitat can be obtained.
Studies of individuals’ morphometrical parameters have indicated that generative plants are presented by 1–7 shoots (mean 2.1). These had length from 56–131 cm (mean 92.6cm). Vegetative individuals were also counted. These plants were presented by rosettes containing three leaves in average with variation from one to seven leaves (Table 3).
Generative reproduction has been estimated through counting the number of umbels per generative individual, number of umbellules per umbel and number of fruits per umbellule (Table 4). Fruit production of S. silaus individuals was determined using the product of these three parameters. As a result, in studied population mean number of fruits per plant was calculated as 10.0 × 8.2 × 61.2 = 5,018.
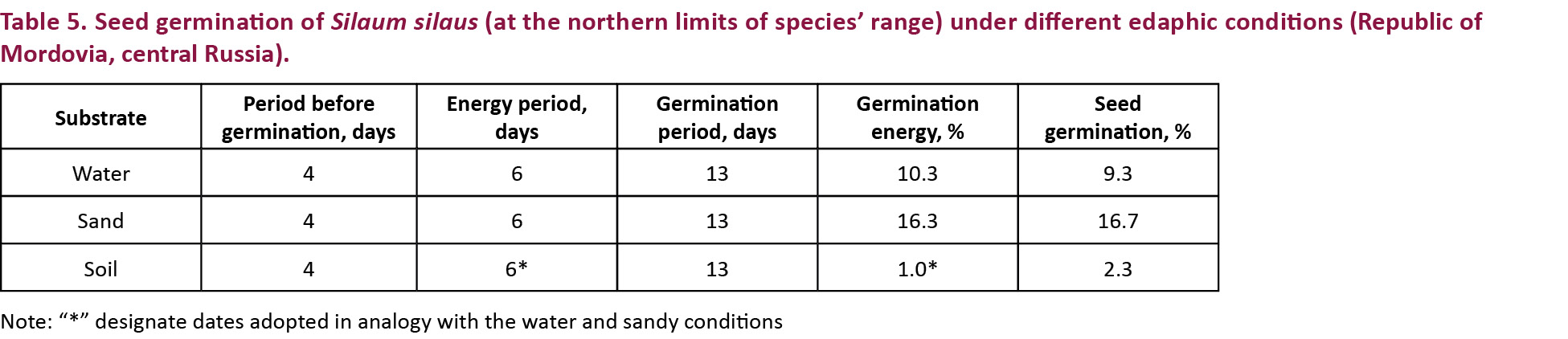
Seed characteristics were determined through counting of seed mass and seed germination. Seed mass was equal to 1.36mg (Table 4). Seed germination was estimated visually by removing germinated seeds out of growth chambers. Seeds started to germinate on the fourth day of the experiment. Germination period lasted 13 days independent of edaphic conditions (Table 5). The minimum period from seed sowing to its germination was three days in all variants of edaphic conditions. This parameter highly varied in different replications or the seeds, however, did not germinate completely. In six replications of the experiment, the period from seed sowing to its germination was three days; in four replications - four days; in four replications - five days; in one replication (in soil conditions) - eight days, and no seed germinated in three replications (in soil conditions).
The maximum number of germinated seeds per day for S. silaus was on day six after beginning of seed germination in both water and sand conditions (Fig. 2). Thus, the energy period of S. silaus seeds was six days. Peak of seed germination values has not been determined in soil conditions (Fig. 2). In this case, the sixth day was used as the period to calculate energy period and germination energy in analogy with water and sand germination conditions. The maximal germination rate of S. silaus seeds was achieved on sand substrate (Table 5).
Discussion
The dominance of hemicryptophytes group in life-form spectrum and sub-mesophytes and sub-xerophytes groups in relation to moisture conditions is considered as typical for similar habitats in forest-steppe zone of Eastern Europe (Lisetskii et al. 2016). Furthermore, a high number of meadow and steppe species indicates affiliation of studied habitat to the northern meadow steppes. Highest species richness in the weed group indicates the significant disturbance of vegetation cover (e.g., Lososová et al. 2006). This is caused by grazing pressure in the last years (Silaeva et al. 2008; 2010–2016 pers. obs.). At present, grazing is less intense due to decrease in cattle population by more than 50% (Pavlov 2013). This may be the reason why S. silaus occupation has increased more than 12 times (from 800–9876 m2) over the last 15 years. Also, populations’ vitality of other regionally rare species (e.g., Echinops ritro L., Stipa tirsa Steven) has also increased in this location (2016 pers. obs.), as it was shown for another steppe plant, Scutellaria baicalensis Georgi (Sandanov et al. 2017).
Interpretation of environmental factors’ values in the studied habitat, obtained using phytoindication method has allowed the brief description of S. silaus habitat according to Tsyganov’s (1983) classification. According to climatic scales, the studied habitat is ranked between values typical for habitats in sub-boreal and nemoral termozones with tendency towards the latter. It belongs to the mainland climate group in sub-arid (water balance (difference between amounts of precipitation and evapotranspiration) is about 100mm per year) conditions of temperate winters (the lowest temperature of coldest month is equal to –120С). The latter result is close to that presented in recent publications on climate of Mordovia (Yamashkin 2012; Bayanov 2015). According to edaphic scales, S. silaus grows on sub-acidophilous (pH ≈ 6.6), nitrogen-poor soils with sub-eutrophic salt regime (salt availability ≈ 200mg/l) under weak to moderately variable humidification. In relation to water content, this habitat has sub-meadow water regime of soils. It occurs when water income (outflow) exceeds precipitation; e.g., the transit surface runoff at slopes. Ground waters lie at a depth of up to 8–10 m. According to coenotic scale (shading), the studied location falls between groups of open and semi-open habitats. Low variation of environment factors’ values may be explained by proximity of sampled plots.
According to Bischoff (2002), the maximum number of S. silaus seedlings is observed within a radius of 1.5m around the mother plant. This seems to be related with limiting values of shoot height, obtained in S. silaus population studied in Mordovia (Table 3). Therefore special investigations of this issue are needed in habitats with different abundance degrees of S. silaus.
Fruit production (5,018 fruits per plant) is much higher than the 1,067 to 2,077 fruits per plant in the Saale River floodplain near Holleben (Bischoff 2000). The large number of fruits in the population studied may represent an adaptation to unfavourable conditions under grazing pressure at the studied locality. Large numbers of small seeds and their dispersal by livestock could be a reason driving expansion of the S. silaus’ population. This is indirectly confirmed by the conclusion that flooding, mowing machinery and grazing animals may disperse seeds over much longer distances (Bischoff et al. 2009). But they are efficient only if they coincide with large seed set and act in the right direction. The latter conditions are consistent with the data obtained on S. silaus seed mass. It has amounted to 1.36mg in Mordovian population, while seeds from flood-meadows in the northern Upper Rhine River (Germany) had mass of 2.43mg (Hölzel & Otte 2004). This may be another reason driving the population to expand over the last 15 years. An almost twofold difference of this parameter cannot be explained only by environment conditions due to their similarity. Perhaps, it is explained by peripheral location of studied S. silaus population. Additional studies are needed to understand this phenomenon.
Seed germination study in experimental conditions has demonstrated low germination rates (from 2.3% on soil to 16.7% on sand) of seeds from Mordovia, while this parameter has varied from 26.8–66.6 % in field conditions in the Saale River floodplain near Holleben (Bischoff 2000). Further studies of seed germination ecology are needed to understand the environmental preferences of S. silaus seeds. Furthermore, it was found that seeds of the endangered plant germinated best on sand substrate. Most likely, this result is not associated with ecological preferences of this floodplain meadow species. That is rather an indication that sand is the most appropriate substrate for seed germination in relation to any plant as it was shown in experiments with both alien and native species (e.g., Gladunova et al. 2014, 2016; Dutra et al. 2016; Radivojevic et al. 2016).
Conclusions
The results showed that the S. silaus population at the northern limit of its range inhabits the floodplain meadow community. According to observations over 15 years, significant disturbance of vegetation cover caused by grazing impact does not limit population vitality. On the contrary, the area occupied by the population had increased by over 12 times over this period. Small size and large number of seeds, as well as moderate levels of livestock grazing may be key reasons behind this change.
Perhaps, such prosperous status of the plant population at the limit of the species’ range may be explained by changing of K-strategy to the r-strategy according to MacArthur & Wilson (1967) concept. Our results on the increase of S. silaus seeds’ number together with their smaller mass may be considered as a support of this view. Similar results are true for some invasive plant species in their introduced range (e.g., Gladunova et al. 2014). Despite high seed number, their germination rate and the germination energy were very low irrespective of edaphic conditions. It is interesting that seed germination was very low in rough substrate (soil), although this substrate should be typical for S. silaus in floodplain meadows in contrast to other germination media (water, sand).
Additional studies on the population of S. silaus in different parts of its range are needed. Particularly, population-based studies would be appropriate and indicative in saline steppes of closely located territories (e.g., Penza region) where S. silaus occurs in high abundance.
References
Abeli, T., R. Gentili, A. Mondoni, S. Orsenigo & G. Rossi (2014). Effects of marginality on plant population performance. Journal of Biogeography 41: 239–249; http://doi.org/10.1111/jbi.12215
Bayanov, N.G. (2015). Climate changes of the northwest of Mordovia during the period of existence of the Mordovia Reserve according to the meteorological observations in Temnikov. Proceedings of the Mordovia State Nature Reserve 14: 212–219. (in Russian)
Beatty, G.E., P.M. McEvoy, O. Sweeney & J. Provan (2008). Range-edge effects promote clonal growth in peripheral populations of the one-sided wintergreen Orthilla secunda. Diversity and Distribution 14: 546–555; http://doi.org/10.1111/j.1472-4642.2008.00469.x
Birkeland, S., I.E.B. Skjetne, A.K. Brysting, R. Elven & I.G. Alsos (2017). Living on the edge: conservation genetics of seven thermophilous plant species in a high Arctic archipelago. AoB PLANTS 9: plx001; http://doi.org/10.1093/aobpla/plx001
Bischoff, A. (2000). Dispersal and re-establishment of Silaum silaus (L.) in floodplain grassland. Basic and Applied Ecology 1: 125–131; http://doi.org/10.1078/1439-1791-00018
Bischoff, A. (2002). Dispersal and establishment of floodplain grassland species as limiting factors in restoration. Biological Conservation 104: 25–33.
Bischoff, A., G. Warthemann & S. Klotz (2009). Succession of floodplain grasslands following reduction in land use intensity: the importance of environmental conditions, management and dispersal. Journal of Applied Ecology 46: 241–249; http://doi.org/10.1111/j.1365-2664.2008.01581.x
Borovik, L.P. (2011). Peculiarities of segetal plant communities of the east of Lugansk region as an initial stage of the abandoned fields succession. The Journal of V.N. Karazin Kharkiv National University. Series biology 14: 33–41. (in Russian)
Buzuk, G.N. & O.V. Sozinov (2009). Regression analysis in phytoindication (on example of D.N. Tsyganov’s ecological scales). Botany (Researches) 37: 356–362. Minsk: Pravo i ekonomika. (in Russian).
Donath, T.W., N. Hölzel & A. Otte (2006). Influence of competition by sown grass, disturbance and litter on recruitment of rare flood-meadow species. Biological Conservation 130: 315–323; http://doi.org/10.1016/j.biocon.2005.12.022
Dutra, F.A., M.M. Araujo, D.G. Rorato & P. Mieth (2016). Seed germination and seedling emergence of Luehea divaricata Mart. et. Zucc. in different substrates. Ciência Florestal 26: 411–418; http://doi.org/10.5902/1980509822744
Fröberg, L. (2009). Silaum Mill. In: Flora Nordica electronic version code 6b. http://www.floranordica.org/Review/-Review_public/accounts/Silaum.html Accessed March 2017.
Gladunova, N.V., E.V. Vargot & A.A. Khapugin (2014). Oenothera biennis L. (Onagraceae) in the Republic of Mordovia (Russia). Russian Journal of Biological Invasions 5: 12–17; http://doi.org/10.1134/S2075111714010044
Gladunova, N.V., A.A. Khapugin & E.V. Vargot (2016). Bidens frondosa L. (Asteraceae) in the Republic of Mordovia (Russia). Russian Journal of Biological Invasions 7: 129–136; http://doi.org/10.1134/S2075111716020053
Guardiola, M., X. Oliver & J.F. García (2014). Les poblacions de Silaum silaus (Umbelliferae) a Catalunya: distribució, ecologia, amenaces i estatus. Orsis: organismes i sistemes 28: 55–73. (in Catalan)
Hammer, Ø., D.A.T. Harper & P.D. Ryan (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4: 9.
Hölzel, N. & A. Otte (2004). Ecological significance of seed germination characteristics in flood-meadow species. Flora 199: 12–24.
IUCN (2012a). IUCN Red List Categories and Criteria: Version 3.1, second ed. Switzerland and Cambridge, Gland, UK, 32pp.
IUCN (2012b). Guidelines for Application of IUCN Red List Criteria at Regional and National Levels: Version 4.0. Switzerland and Cambridge, Gland, UK, 41pp.
IUCN (2014). IUCN Standards and Petitions Subcommittee. Guidelines for Using the IUCN Red List Categories and Criteria. Version 11. Prepared by the Standards and Petitions Subcommittee. Retrieved 02 February 2017 from http://www.iucnredlist.org/documents/RedListGuidelines.pdf.
Khapugin, A.A. & G.G. Chugunov (2015). Two populations of Lunaria rediviva L. (Cruciferae Juss.) at the eastern edge of its range. Wulfenia 22: 83–94.
Khapugin, A.A., T.B. Silaeva, E.V. Vargot, G.G. Chugunov, G.A. Grishutkina, O.G. Grishutkin, E.V. Pismarkina & J.S. Orlova (2017a). Estimation of taxa included in the first volume of the Red Data Book of the Republic of Mordovia (Russia) using the IUCN Red List Categories and Criteria. Nature Conservation Research 2(Suppl. 1): 164–189. (in Russian); http://doi.org/10.24189/ncr.2017.004
Khapugin, A.A., G.G. Chugunov, E.V. Vargot & T.B. Silaeva (2017b). Vascular plants at the protected areas network of the Republic of Mordovia: present status and prospects, pp. 203–231. In: Mukul, S.A. & A.Z.M.M. Rashid (eds.). Protected Areas: Policies, Management and Future Directions. Nova Science Publishers, Inc., USA, 239pp.
Lisetskii, F.N., B. Sudnik-Wojcikowska & I.I. Moysiyenko (2016). Flora differentiation among local ecotopes in the transzonal study of forest-steppe and steppe mounds. Biology Bulletin 43: 169–176; http://doi.org/10.1134/S1062359016010106
Lososová, Z., M. Chytrý, I. Kühn, O. Hájek, V. Horáková, P. Pyšek & L. Tichý (2006). Patterns of plant traits in annual vegetation of man-made habitats in central Europe. Perspectives in Plant Ecology, Evolution and Systematics 8(2): 69–81; https://doi.org/10.1016/j.ppees.2006.07.001
MacArthur, R.H. & E.O. Wilson (1967). The Theory of Island Biogeography. Princeton University Press, Princeton, 203pp.
Maevsky, P.F. (2014). Flora of Center of European part of Russia. KMK Scientific Press Ltd., Moscow, 635pp. (in Russian)
Menglan, S. & M.F. Watson (2005). 65. Silaum Mill., pp. 134–135. In: Zhengyi, W., P.H. Raven & H. Deyuan (eds.). Flora of China. Missouri Botanical Garden, St. Louis, MO: Science, Beijing.
Naik, M.C., M.A. Kumar & B.R.P. Rao (2016). On the discovery of Dimeria hohenackeri (Poaceae) from the Andaman Islands, a hitherto known endemic and endangered grass species of southwestern peninsular India. Journal of Threatened Taxa 8(14): 9678–9680; http://doi.org/10.11609/jot.2500.8.14.9678-9680
Nikolaeva, M.G., M.V. Razumova & V.N. Gladkova (1985). The Handbook on Germination of Resting Seeds. Nauka, Leningrad, 348pp. (in Russian)
Novikova, L.A. & D.V. Pankina (2013). Characteristic of the vegetation saline plots in Malaja Serdoba and Serdobsk districts of Penza region. Mordovia University Bulletin 3–4: 21–26. (in Russian)
Pavlov, M.Y. (2013). Dynamics of a livestock of cattle in the regions of Russia depending on the share of livestock in farms. Scientific Notes of the Bauman Kazan State Academy of Veterinary Medicine 2: 313–317. (in Russian)
Pimenov, M.G. & T.A. Ostroumova (2012). Umbelliferae of Russia. KMK Scientific Press Ltd., Moscow, 484pp. (in Russian)
Pimenov, M.G. & V.N. Tikhomirov (1979). Typificatio generum Umbelliferarum florae URSS. Novosti sistematiki vysšich rastenij 16: 154–166. (in Russian)
Radivojevic, L., M. Saric-Krsmanovic, J.G. Umiljendic, D. Bozic & L. Santric (2016). The Impacts of temperature, soil type and soil herbicides on seed germination and early establishment of Common Milkweed (Asclepias syriaca L.). Notulae Botanicae Horti Agrobotanici Cluj-Napoca 44: 291–295; http://doi.org/10.15835/nbha44110265
Raunkiaer, C. (1934). The Life Forms of Plant and Statistical Plant Geography. Clarendon Press, Oxford, 632pp.
Sandanov D.V., B.B. Naidanov & V.M. Shishmarev (2017). Influence of regional and local environmental factors on the distribution and population structure of Scutellaria baicalensis Georgi. Tomsk State University Journal of Biology 38: 89–103. (in Russian); http://doi.org/10.17223/19988591/38/5
Shennikov, A.P. (1950). Ecology of Plants. Sovetskaya Nauka, Moscow, 375pp. (in Russian)
Silaeva, T.B. & N.A. Barmin (1995). Floristic records in Mordovia, pp. 66–67. In: Tikhomirov, V.N. (ed.). Floristic Studies in Central Russia. Lipetsk, 156pp. (in Russian)
Silaeva, T.B., I.V. Kiryukhin, E.V. Pismarkina, G.G. Chugunov, E.V. Vargot, A.M. Ageeva, V.M. Smirnov & A.A. Khapugin (2008). Rare Plants and Fungi: Materials for Maintenance of the Red Book of the Republic of Mordovia for 2008. Publisher of the Mordovia State University, Saransk, 102pp. (in Russian)
Silaeva, T.B., I.V. Kiryukhin, G.G. Chugunov, V.K. Levin, S.R. Mayorov, E.V. Pismarkina, A.M. Ageeva & E.V. Vargot (2010). Vascular Plants of the Republic of Mordovia (Synopsis of Flora). Publisher of the Mordovia State University, Saransk, 352pp. (in Russian)
Sotek, Z. (2007). Life history of Plantago coronopus L. at the limit of its range. Ekológia (Bratislava) 26: 14–29.
Srivastava, A., Y.P. Sharma, O.P.S. Vidyarthi & S.K. Srivastava (2017). Conservation status assessment and new population record of the threatened Golden Himalayan Spike Phlomoides superba (Royle ex Benth.) Kamelin & Makhm. from Jammu & Kashmir, India. Journal of Threatened Taxa 9(4): 10089–10095; http://doi.org/10.11609/jott.2875.9.4.10089-10095
The Plant List (2013). Version 1.1. Retrieved 12 June 2017 from: http://www.theplantlist.org/
Tikhomirov, V.N. (2014). Silaum Mill., pp. 293. In: Flora of Middle Belt of European Part of Russia, vol. 11. KMK Scientific Press Ltd., Moscow, 635pp. (in Russian)
Tsyganov, D.N. (1983). Phytoindication of Ecological Regimes in the Mixed Coniferous-Broad-Leaved Forest Subzone. Nauka, Moscow, 197pp. (in Russian).
Tutin, T.G. (1968). Silaum Miller., pp. 342. In: Tutin T.G., V.H. Heywood, N.A. Burges, D.M. Moore, D.H. Valentine, S.M. Walters & D.A. Webb (eds.). Flora Europaea - Vol. 2. Cambridge University Press, London, 499pp.
Willner, W., N. Sauberer, M. Staudinger & L. Schratt-Ehrendorfer (2013). Syntaxonomic revision of the Pannonian grasslands of Austria - Part I: Introduction and general overview. Tuexenia 33: 399–420.
Wróblewska, A. & E. Brzosko (2006). The genetic structure of the steppe plant Iris aphylla L. at the northern limit of its geographical range. Botanical Journal of the Linnean Society 152: 245–255; http://doi.org/10.1111/j.1095-8339.2006.00568.x
Yamashkin, A.A. (ed.). (2012). Geographical Atlas of Republic of Mordovia. Publisher of the Mordovia State University, Saransk, 204pp. (in Russian)