Article
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 March 2017 | 9(3): 9892–9902

A comparative study of the diet of the Indian Eagle Owl Bubo bengalensis (Franklin, 1831) from two distinct habitats in the Tamil Nadu - Puducherry area, southern India
M. Eric Ramanujam 1 & Tushita Singh 2
1 Pitchandikulam Bioresource Centre / Pitchandikulam Forest Consultants, Auroville, Tamil Nadu 605101, India
2 37-A,1st Floor, Goubert Padasalai Street, Vazhaikulam, Pondicherry 605001, India
1 ericramanujamowl@yahoo.com (corresponding author), 2 tushitasingh87@gmail.com
doi: http://doi.org/10.11609/jott.2438.9.3.9892-9902
Editor: Reuven Yosef, Ben Gurion University, Eilat, Israel. Date of publication: 26 March 2017 (online & print)
Manuscript details: Ms # 2438 | Received 23 December 2015 | Final received 11 February 2017 | Finally accepted 25 February 2017
Citation: Ramanujam, M.E. & T. Singh (2017). A comparative study of the diet of the Indian Eagle Owl Bubo bengalensis (Franklin, 1831) from two distinct habitats in the Tamil Nadu - Puducherry area, southern India. Journal of Threatened Taxa 9(3): 9892–9902; http://doi.org/10.11609/jott.2438.9.39892-9902
Copyright: © Ramanujam & Singh 2017. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: None.
Competing interests: The authors declare no competing interests.
Author Details: M. Eric Ramanujam has been a wildlife illustrator for over two decades and has a background in the advertising industry. Since 1997 he has been involved in full time conservation and has been part of a team which undertook surveys of the Kaliveli region near Puducherry, Eastern Ghats and Adyar wetland complex in Chennai. His main sphere of interest is the natural history of the Indian Eagle Owl Bubo bengalensis. Tushita Singh was employed to study rodent population dynamics in agroecosystems, ravines and sacred groves. She has a Masters in Ecology & Environmental Sciences from Department of Ecology & Environmental Sciences, Pondicherry University.
Author Contribution: MER did the analysis and identification of pellet contents and prepared the text. TS did the field work and prepared the final tables and figures.
Acknowledgements: We are thankful to Luke Bowley (alias Govinda), V. Arun, Vijay Sharma and C. Parusuraman for their help in locating nests and collecting pellets at Arunachala. We also wish to thank A. Lakshmikantan for photographing the nests and rodents, as well as Veeramani who supervised the trapping of field rodents with the Irula field assistants, Sekar and Krishnamurthi.
Abstract: A total of 166 prey items of the Indian Eagle Owl Bubo bengalensis were identified from pellets, pellet remains and prey remains—84 from Arunachala Hill and 82 from Pondicherry University campus. The total biomass encountered from pellet and prey remains was 22,620g; of which, 11,241g was from Arunachala and 11,380g was from Pondicherry University. Out of the 166 prey items 102 were non-volant small mammals accounting for a biomass of 13,974g; of which 5,617g (49.94%) was from Arunachala and 8,357g (73.42%) was from Pondicherry University. Even among these, murid rodents dominated with 44.99% in Arunachala and 70.13% in Pondy University. Thirteen birds of three species were predated upon and only partly consumed; hence biomass could not be inferred as the amount ingested was not determinable. Of these 10 were Mlivus migrans, a raptor co-inhabiting the same area. Anurans occurred for a combined biomass of 12.87% in both areas. The rest, viz., Coleoptera, Orthoptera and Paratelphusa sp. accounted for a paltry biomass of 0.51%. In Arunachala the constant food of the owls were the field rodents Millardia meltada (15.47%), Bandicota bengalensis (8.74%) and Tatera indica (11.65%), and a single Bandicota indica (6.07%), which is a species found around rural habitations. The first two species were conspicuous by their absence in Pondicherry University and the Tatera indica was encountered in very small quantities (3.70%). Uniquely enough, the terrestrial and fossorial forms of urban rodents formed the basic food of Bubo bengalensis in Pondicherry University - Rattus rattus (20.64%) and Bandicota indica (43.94%). We have compared non-volant small mammal prey of the regions we studied with those of Maharastra and the results are discussed.
Keywords: Intraguild aggression/predation, Linnaean and Wallacean shortfalls, non-volant small mammals, murid rodents, prey spectrum.
Introduction
The genus Bubo contains the world’s largest species of owls, most, if not all of which, are tertiary consumers and excellent indicators of the ecosystems they inhabit. Twenty-five species belong to this genus (Konig & Weick 2010; Mikkola 2012), of which the prey spectra of nine species are known. Some species have been extensively studied, especially the Holarctic species—Eurasian Eagle Owl Bubo bubo (e.g., Choussy 1971; Blondel & Badan 1976; Mysterud & Dunker 1982; Orsini 1985; Donazar 1987; Simeonov et al. 1998), American Great Horned Owl Bubo virginianus (e.g., Llinas-Gutieerrez et al. 1991; Trejo & Grigera 1998) and Snowy Owl Bubo (Nyctea) scandiaca (e.g., Allan 1977; Boxall & Lein 1982; Robinson & Becker 1986; Potapov & Sale 2012). In fact the stage has been reached where international food spectrums can be compared and analysed (Jaksic & Marti 1984), in addition to influencing predictive models of habitat preference (Martinez 2003) and landscape structure and breeding performance (Penteriani et al. 2002). Food spectrum studies of other owl species of the genus Bubo are not that intensively recorded though four African species have been studied, viz., Pharaoh Eagle Owl Bubo ascalaphus (see Carpentier 1934; Vien & Thevenot 1978; Shehab & Ciach 2008), Spotted Eagle Owl Bubo africanus (see Nel 1969; Demeter 1982; Medelson 1989), Cape Eagle Owl Bubo capensis (see Brooke 1973; Steyn & Myburgh 1983; Allen 1995; Gargett & Grobler 1976; Rodel et al. 2002;) and Milky or Verreaux’s Eagle Owl Bubo lacteus (see Brown 1965), and in India work on only one species has been carried out, Indian or Rock Eagle Owl Bubo bengalensis (see Ramanujam 2006, 2015; Pande & Dahanukar 2011; Pande et al. 2011).
Bubo bengalensis is partial to rocky low hills and ravines, but is also found in semi-desert, deciduous, disturbed or secondary woodland, and even orchards or near villages. It roosts in rock cavities, and sits on rock stacks or ledges and hunts mammals and other prey, including large insects, using sit-and-wait methods or during flight (Rasmussen & Anderton 2012). In India, prey spectrums of the species have been derived from the states of Tamil Nadu - Puducherry in southern India (Ramanujam 2006, 2015) and in Maharastra in central India (Pande & Dahanukar 2011; Pande et al. 2011; Patki et al. 2014) and it is now possible to compare the prime prey base which comprises non-volant small mammals, especially murid rodents, between sites. In addition this paper attempts to add two more sites where pellets and prey remains have been collected during the nesting season in Tamil Nadu and Puducherry which could further enhance the knowledge of the prey base of the species in relation to the difference in habitat types between the two areas.
Materials and Methods
Study areas
Arunachala Hill (12011’N & 79002’E), also known as Arunagiri, Arunachalam, Arunai, Sonagiri and Sonachalam, is a holy hill at Tiruvanamalai in Tamil Nadu (Image 1). It is an imposing landmark with an elevation of nearly 800m. It is an assemblage of charnockite and associated rocks older than 3,500 million years (www.arunachala-live.com). It is surrounded by fields, primarily rice and sugarcane.
Pondicherry University (12001’N & 79085’E) lies in Kalapet on the northern outskirts of the Union Territory of Puducherry (formerly Pondicherry, a part of French India) and comprises an area of 780acres (Image 1). Puducherry is not a contiguous area and many parts alternate between Tamil Nadu and the Union Territorry. For example if one drives down from Chennai via the East Coast Road, one will first touch Kalapet, then travel through the Auroville area in Tamil Nadu and again touch Puducherry City at Muthialpettai. In addition, there are isolated pockets like Sanjeevnagar in Auroville surrounded by Tamil Nadu on all sides. The area between Auroville and Kalapet is characterized by a number of ravines and gullies that have been created by surface and rill erosion during the monsoons when runoff water is carried into the Bay of Bengal. The ravines of Pondicherry University fall in these no-man’s lands and hence have received little attention from the authorities. With a student capacity of around 10,000, the built up area is considerable with 19 hostels, 11 department blocks and canteens and cafeterias. As a result all the raw sewage, solid waste and left over food find their way into the ravines (Seshadri 2013) and these attract rodent vermin like in cities, hence the area can be considered to be an urban ravine ecosystem like the ravines adjoining the Jawaharlal Nehru Institute of Postgraduate Medical Education and Research (JIPMER). For Locations refer to Image 1.
Methods
The pellet analysis method has been the fundamental way of analyzing prey spectrums of owls and has withstood the test of time (Errington 1930, 1932; Fischter 1941). Of late this method has come under criticism due to biases, and it has been suggested that a combination of different methods may yield more accurate estimates of overall diet (Rosenberg & Cooper 1990; Simmons et al. 1991; Marchesi & Pedrini 2002). The latter was the methodology we adopted as we searched for pellets, pellet remains and prey remains at the nests / vicinity of nests in the study area (and in one earlier instance at Nanmangalam Reserve Forest in Chennai relied on direct observation). On the Arunachala Hill a nest in a fissure of rock with three young was discovered in March 2015 (Images 2 & 3). When the young left the nest we were able to collect 48 entire pellets and 21 broken up pieces. In the ravines of the Pondicherry University campus a disused nest was discovered in July 2015, which contained 45 broken up and 18 entire pellets. The summer months of this year had unprecedented rains and it is quite possible that some pellets / pellet remains were washed away. In addition prey remains of 13 birds, all partly consumed, were encountered, but no mammal remains were in evidence in spite of us scouring the area thoroughly. The pellets and pellet remains were bagged and kept in a bake oven for 24 hours. The pellets were then subjected to NaOH treatment and the contents were separated and subjected to taxonomical identification. Mammal remains were identified relying on established literature (Ramanujam 2004; Talmale & Pradhan 2009), birds from their remains and arthropods according to Mani (1990), in addition to cross checking with our study collection and those now housed in the repository of Wildlife Information and Liaison Development (WILD) in Coimbatore. Biomass was derived from the mean weight of 25 individuals collected by Irula tribals in fields (Image 4), human habitations and ravines and weighed using scales accurate to 0.05 g. Biomass was chosen as the principal measure to assess intake of food as it gives a fair idea about the quantity of food consumed by the owls.
The categories of food components consumed was assessed according to a pre-established system for calculating trophic connections of owls in India (Kumar 1985; Verzhutskii & Ramanujam 2002; Ramanujam & Verzhutskii, 2004; Ramanujam 2006, 2015). This recognizes five classes: Basic food - prey occurring with a frequency above 20%; Constant food - prey occurring with a frequency between 5–20 %; Constant food – prey occurring with a frequency between 5–20 %; Supplementary food - prey occurring with a frequency between 1–5 %; and Occasional food - prey occurring with a frequency below 1%.


Results
A total of 166 prey items were identified from pellets and prey remains - 84 from Arunachala Hill and 82 from Pondicherry University campus (Table 1). The total biomass consumed was 22,620g, out of which 11,241g was from Arunachala Hill and 11,380g was from Pondicherry University (Figs. 1 & 3). Out of the 166 prey items 102 items were non-volant small mammals accounting for a total biomass of 13,974g; out of which 5,617g from Arunachala Hill (49.94%) and 8,357g from Pondicherry University (73.42%) (Figs. 2 & 4). Of the non-volant small mammals murid rodents accounted for a biomass of 44.99% in Arunachala and 70.13% in Pondicherry University. This shows that non-volant small mammals were the bulk of food consumed by Bubo bengalensis in both areas with murid rodents dominating. The presence of the Black Kite Milvus migrans was the single largest biomass in Arunachala Hill (4,397g or 39.11%) and accounted for a sizeable weight in Pondicherry University also (1,885g or 16.56%), and other birds too accounted for a combined biomass of 693g (3.06%) in both areas. Though the biomass predated upon is clear from references to literature (Kimya et al. 1989; Viega & Hiraldo 1990), the biomass ingested was unclear since the carcasses were the worse for wear due to exposure to the elements - most were dehydrated remains and field weights could not be considered for evaluation. This may appear to bias the results, but as only parts of these birds were consumed, it was not reasonable to include these in biomass calculations and assign the category of prey under which they fall. It is a known fact that the pellet and prey remains analysis methods overestimate avian and large prey predatorship (Rosenberg & Cooper 1990; Marchesi & Pedrini 2002), hence corrective factors should be mentioned in any study dealing with raptor diets, especially if biomass is the key factor for analysis (Simmons et al. 1991). Anurans accounted for a combined biomass of 1,610g – 910g (8.09%) at Arunachala and 700g (6.15%) at Pondicherry University. The rest of the prey, which included beetles (Coleoptera), grasshoppers and crickets (Orthoptera) and freshwater or field crabs (Paratelphusa sp.), were of minor importance since they accounted for a total biomass of only 60.56g (0.51%). For further details refer Table 1, and Figs. 1 & 2, for both numerical and biomass percentages.
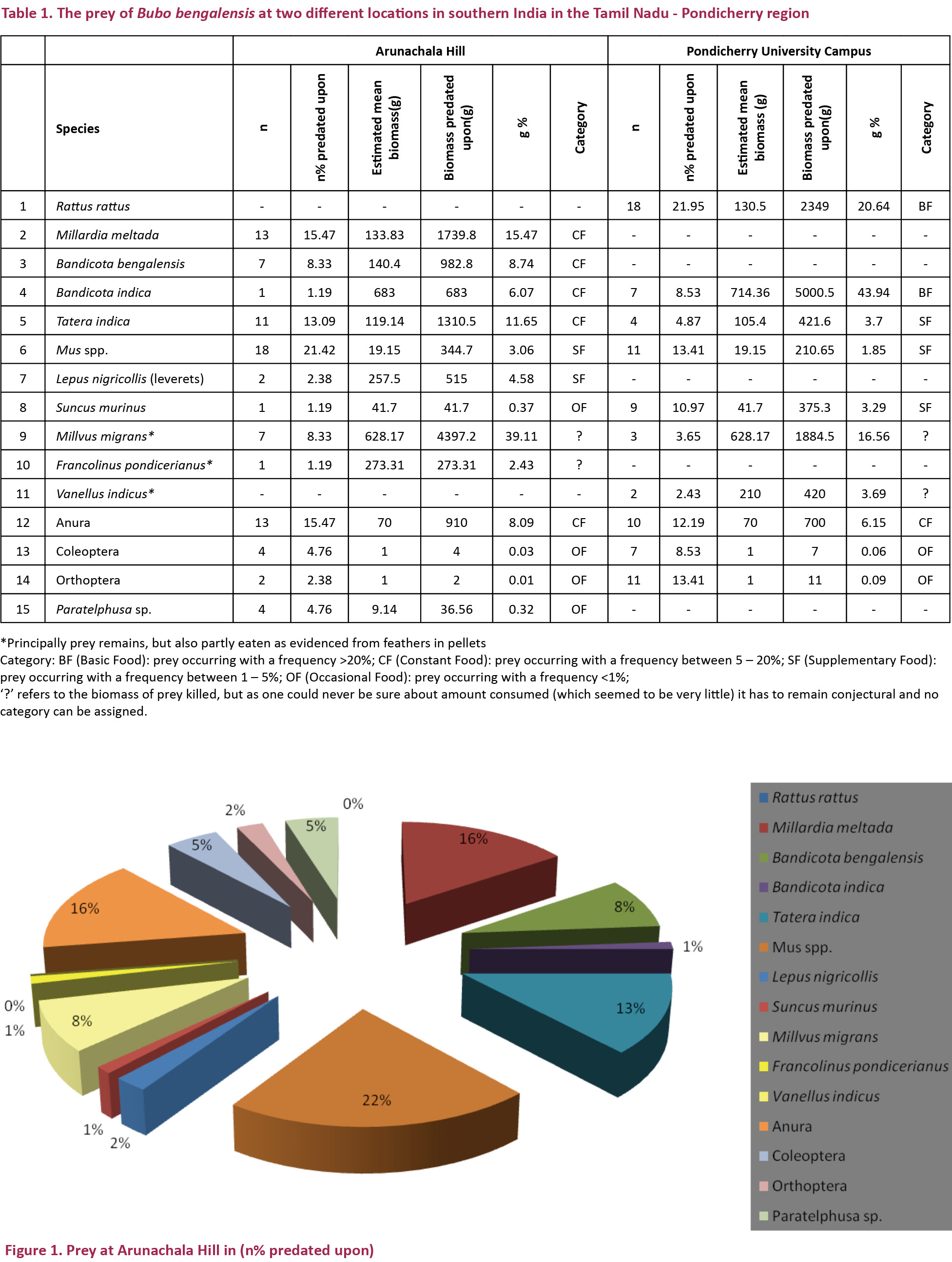
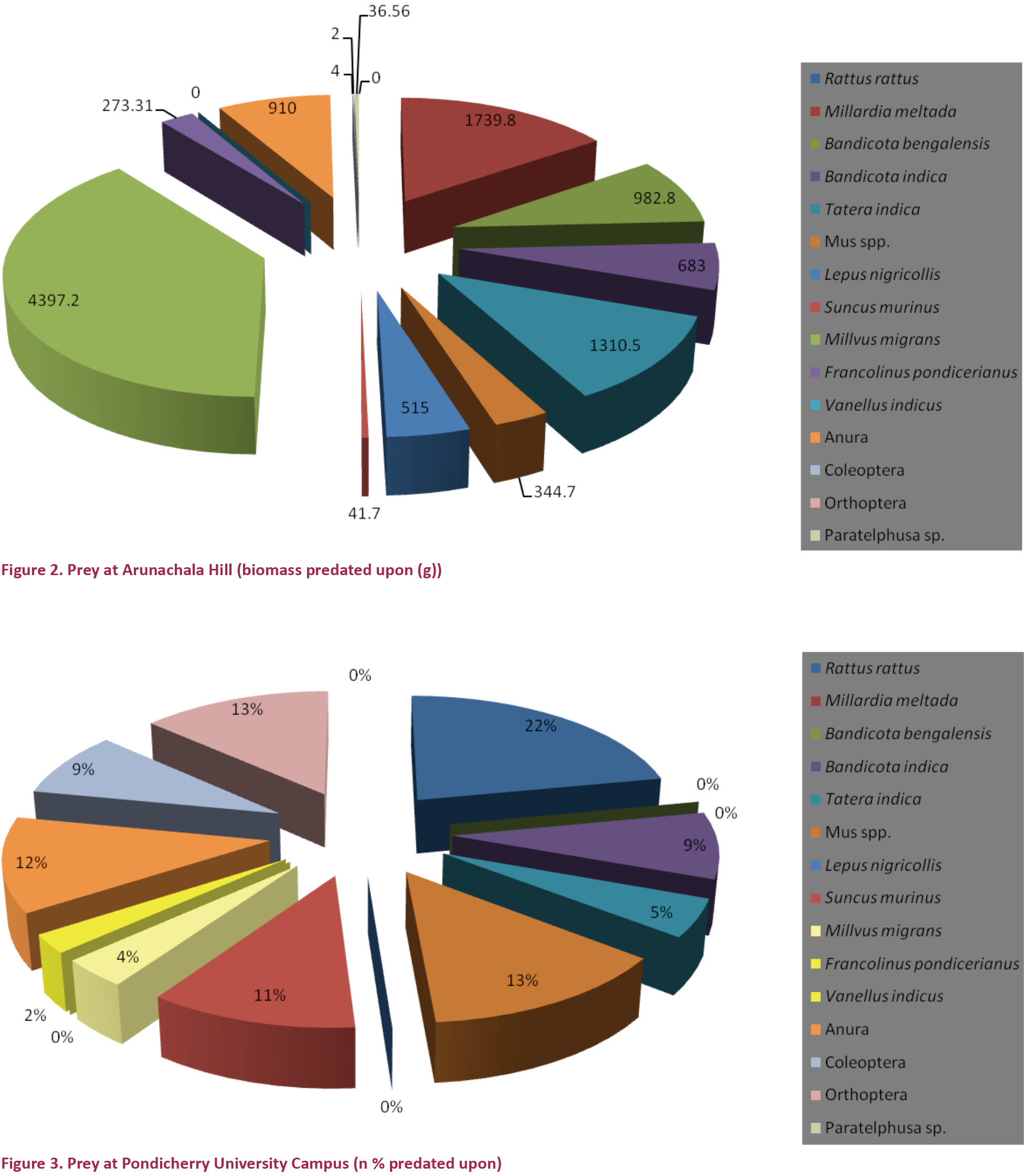
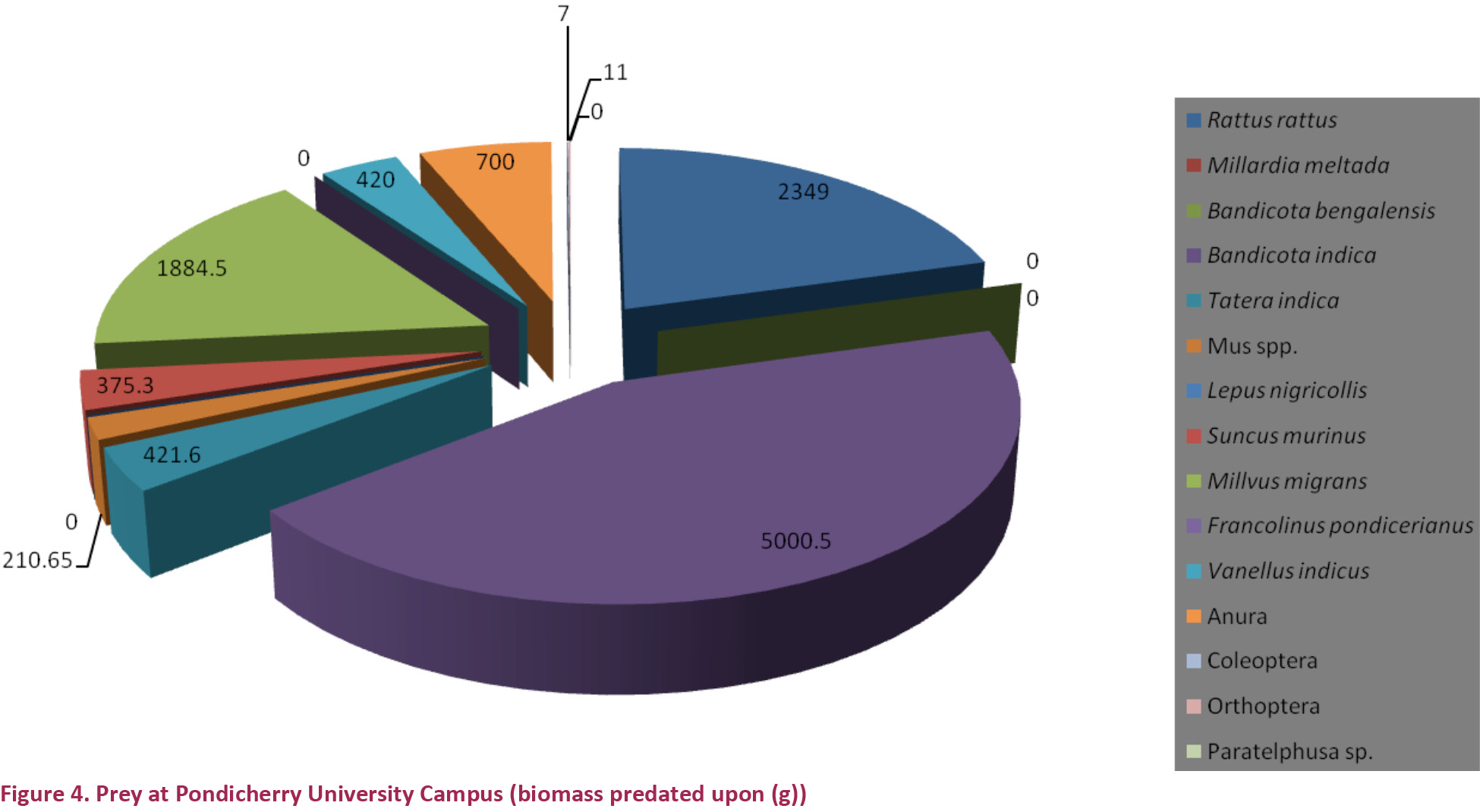
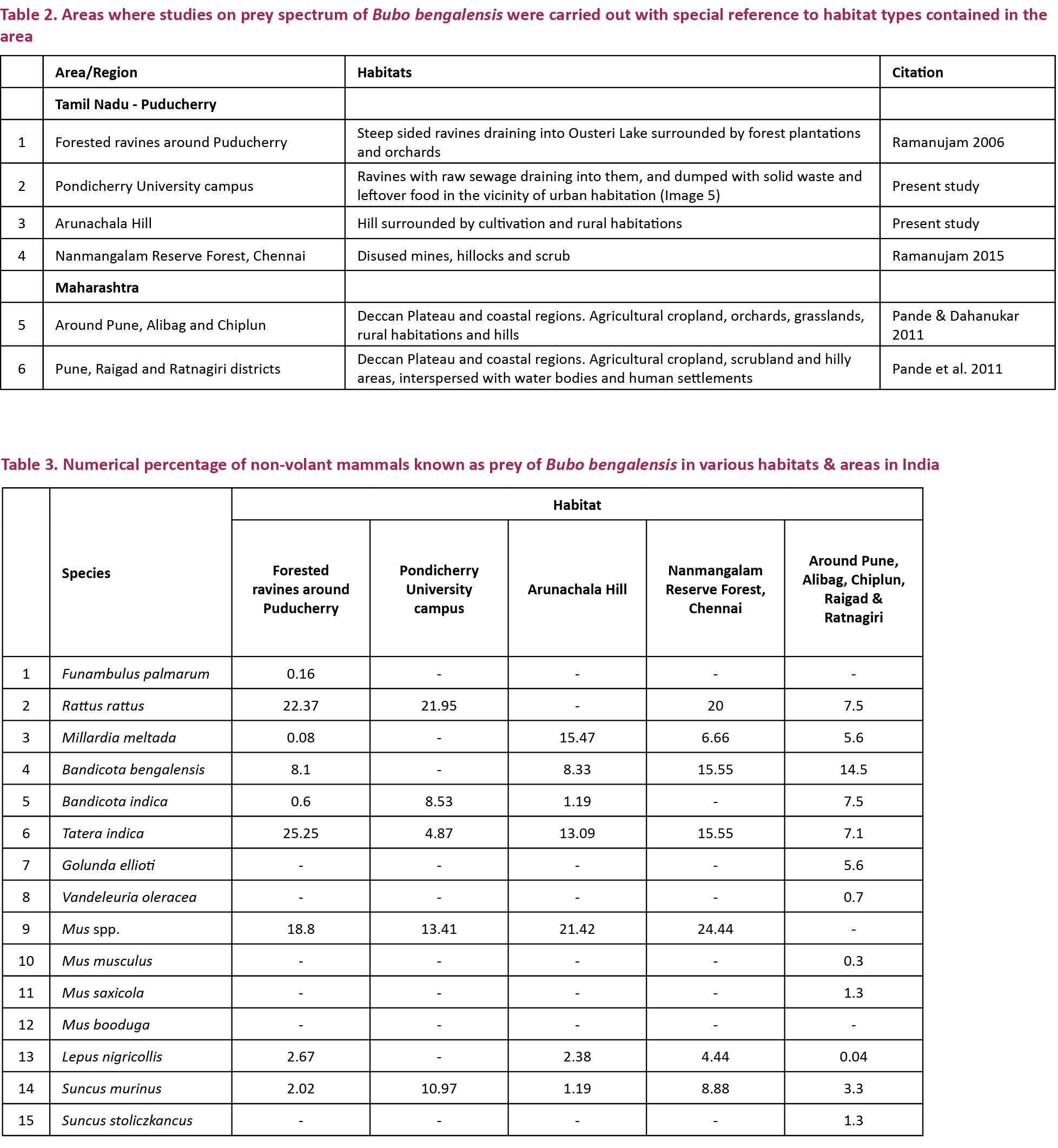

Discussion
It was interesting to note the difference and quantities consumed of non-volant small mammals, especially murid rodents, between the two types of habitats. At Arunachala the constant food (which was taken to be prey items occurring with a frequency of 5–20 %) was the Soft-furred Field Rat Millardia meltada, Lesser Bandicoot or Mole Rat Bandicota bengalensis, Large Bandicoot Rat Bandicota indica and Indian Gerbil Tatera indica - all species commonly found in agricultural lands and rural habitations rather than on hillocks. This discloses the fact that Bubo bengalensis is hunting predominantly in fields surrounding the hillock and this behaviour was attested to from the hillocks near Chingelpet (Romulus Whitaker pers. comm. 2014). In Pondicherry University the first two species were conspicuous by their absence and Tatera indica occurred only as supplementary food (1–5 %). Another feature was that the House Rat Rattus rattus was not encountered in Arunachala since the rufous form is an arboreal forest dwelling species and does not occur in fields, but the dark terrestrial form commonly found in urban areas comprised the basic food along with the Large Bandicoot Rat Bandicota indica in Pondicherry University campus (basic food was taken to be prey items occurring with a frequency of >20%). Seven Bandicota indica accounted for a biomass of 5,000.52g or 43.94%! Both the latter species are commensals of humans and occur in pestilential quantities in cities and around human habitations. The specific identity of mice (Mus spp.) could not be verified because in spite of valuable work having been done in the Indo-Malayan region (Marshall 1977, 1986) many anomalies remain unresolved (Agrawal 2000; Srinivasulu & Pradhan 2003). That the Mus species complex remains unresolved is clearly indicated from the studies of Patki et al. (2014) as even Talmale who was the co-author of “Identification of some small mammal species through Owl Pellet Analysis” could not specifically identify many specimens and grouped them under the heading “Mus species”. Tissue samples of specimens captured by us along with skins and skeletal matter are in the repository of WILD awaiting molecular analysis and it is hoped some light will be shed on the species complex in the near future. Among non-murid small mammals, leverets of the Indian Hare Lepus nigricollis were found in Arunachala and the Musk Shrew Suncus murinus was predated in both habitats, though many more were taken in the Pondicherry University campus probably because they too are commensals of humans.
The presence of partly consumed Black Kite Milvus migrans remains around the nests of Bubo bengalensis may be surprising to some but Eagle Owls are intolerant to the presence of other avian predators, and territorial behaviour and intraguild aggression / predation by Eagle Owls on other owls and diurnal raptors is well established (Mikkola 1976; Real & Manosa 1990; Tella & Manosa 1993; Sergio et al. 2003; Cui et al. 2008; Sergio & Hiraldo 2008). Milvus migrans is often solely thought of as a scavenger, especially of urban areas, but whenever the need presents itself it becomes an active predator (Viega & Hiraldo 1990; Sergio & Boto 1999; Villafuerte & Vinuela 2006). Further to this Milvus migrans is known to share the same nesting areas (Sergio & Pedrini 2003) hence it belongs to the same guild due to overlap of both prey base and habitat space during the breeding season and hence it is not uncommon for it to be attacked by Eagle Owls.
We have determined the prey spectrum and differences in food types, especially concerning non-volant small mammals, between the two habitats but it may be more illuminating to compare the results derived in Tamil Nadu - Puducherry to Maharastra which is the only other state where studies on Bubo bengalensis has been undertaken. Table 2 lists the areas of studies, habitats contained in the study areas and the authors of these studies, while Table 3 elucidates the numerical percentage of non-volant mammals known to occur in the diet of Bubo bengalensis in these areas. We have chosen to ignore other prey types known to be consumed by Bubo bengalensis as the percentage of these were comparatively very low and varied, and this fact has been borne out by studies concerning Bubo bubo in other parts of Asia - for example, in Mongolia (Tumurbat et al. 2009), Iran (Obuch 2014) and Syria (Shehab 2004).
The presence of the diurnal Three-striped Palm Squirrel Funambulus palmarum has been recorded in forested ravines around Puducherry, albeit in low quantity (0.16%). This may be that in undisturbed areas like these Bubo bengalensis is partly diurnal and other large owls do not seem to be strictly crepuscular or nocturnal - the Forest or Spot-bellied Eagle Owl Bubo nipalensis has twice been seen predating on Giant Squirrels Ratufa indica during the day (Kannan 1994, 1998). Rattus rattus seems to be the basic food (>20%) in southern India, except for Arunachala where it is conspicuous by its absence. In comparison the rat forms the constant food (5–20 %) in Maharashtra. Millardia meltada seems to be an anomaly: it seems to prefer agricultural fields in southern India (15.47%) and hilly, forested tracts in Maharashtra (35.29% at Melghat) as well as in fields (16.66% at Rahatgaon). Though this may sound contradictory, it is an established fact that it is a species primarily found in rocky areas (Srinivasulu et al. 2004) as well as agricultural fields (Barnett & Prakash 1975; Fall 1977). Bandicota bengalensis is a species found in scrubland (Prakash et al. 1995) and is the reason why it forms the basic and constant food of B. bengalensis in Melghat (23.53%) and Nanmangalam R.F. (15.55%). Even in fields it prefers the wooded / scrub borders (Author pers. obs. & Sanjay Molur pers. comm. August 2012) and hence is found in lower quantities in these biomes. In southern India Tatera indica is found in both forested areas and fields (13.09–25.25 %) but occurs in very low numbers around urban settlements like Pondicherry University (4.87%). In Maharashtra it is poorly represented in the diet of B. bengalensis. The Bush Rat Golunda ellioti and Long-tailed Tree Mouse Vandeleuria oleracea were not encountered in the diet of B. bengalensis in southern India and in very low quantities in Maharashtra. Little can be said about the Mus species complex until some clarity concerning the Linnaean and Wallacean shortfalls emerge. It was surprising that Lepus nigricollis did not feature among the prey of B. bengalensis in Melghat though its absence in an urbanized habitat like Pondicherry University is understandable. Suncus murinus is a species well adapted to all biomes and it is not surprising that it occurs regularly in the diet of owls – in fact it forms the prime prey base of the Barn Owl Tyto alba in certain areas (Mahmood-ul-Hassan et al. 2007). Suncus stoliczkancus is a rare species and it is fortunate that it was found in pellets of B. bengalensis in Maharashtra, which demonstrates that owl pellet analysis is also a useful method for gaining additional insight into small mammal communities and distribution (Choate 1971).
References
Agrawal, V.C. (1967). Taxonomic study of the skulls of Oriental rodents in relation to ecology. Records of the Indian Museum 60: 125 – 326.
Allan, T.A. (1977). Winter food of the snowy owl in northwestern lower Michigan. Jack Pine Warbler 55: 42.
Allen, D. (1995). The diet of the Cape Eagle Owl. Journal of African Raptor Biology 10: 12–27.
Barnett, S.A. & I. Prakash (1975). Rodents of Economic Importance. Arnold Heinmann, New York, 175pp.
Blondel, J. & O. Badan (1976). La biologie du Hibou grand-duc en Provence. Alauda 47: 271–275.
Boxall, P.C. & M.R. Lein (1982). Feeding Ecology of Snowy Owls (Nyctea scandiaca) Wintering in Southern Alberta. Arctic 35: 282–290.
Brooke, R.K. (1973). Notes on the distribution and food of the Cape Eagle Owl in Rhodesia. Ostrich 44: 137–139; http://doi.org/10.1080/00306525.1973.9632627
Brown, L.H. (1965). Observations on Verreaux’s Eagle Owl Bubo lacteus (Temminck) in Kenya. Journal of the East African Natural History Society 25: 101–107.
Carpentier, C.J. (1934). Le pluri-reject quotidian de pelotes par Bubo bubo ascalaphus. Osieaux 4: 353–355.
Choate, J.R. (1971). Notes on geographic distribution and habitats of mammals eaten by owls in southern New England. Transactions of the Kansas Academy of Sciences 74: 212–216.
Choussy, D. (1971). Etude d’une population de grand-ducs Bubo bubo dans le Massif Central. Nois Oiseaux 31: 37–56.
Cui, Q., J. Su & Z. Jiang (2008). Summer diet of two sympatric species of raptors Upland Buzzard (Buteo hemilasius) and Eurasian Eagle Owl (Bubo bubo) in alpine meadow: problem of coexistence. Polish Journal of Ecology 56: 173–179.
Demeter, A. (1982). Prey of the Spotted Eagle Owl Bubo africanus in Awash National Park, Ethiopia. Bonner zoologische Beitrage 33: 283–291.
Donazar, J.A. (1987). Geographic variations in the diet of eagle owls in western Mediterranean Europe. Biology and Conservation of Northern Forest Owl: Symposium Proceedings, 220–224pp.
Errington, P.L. (1930). The pellet analysis method of raptor food habits study. Condor 32: 292 – 296.
Errington, P.L. (1932). Technique of raptor food habits study. Condor 34: 75 – 86.
Fall, M.W. (1977). Rodents in Tropical Rice. Technical Bulletin No. 36, Interpretative Programmes Centre, Denver Wildlife Research Centre, U.S. Fish and Wildlife Service, Colorado.
Fischter, E. (1941). The role of owl pellet analysis in faunistics. Nebraska Bird Review 9: 26–30.
Gargett, V. & J.H. Grobler (1976). Prey of the Cape Eagle Owl Bubo capensis mackinderi Sharpe 1899, in the Matopos, Rhodesia. Arnoldia 8: 1–7.
Jaksic, F.B. & C.D. Marti (1984). Comparitive food habits of Bubo owls in Mediterranean-type ecosystems. The Cooper Ornithological Society 86: 288–296.
Kannan, R. (1994). Forest Eagle Owl (Bubo nipalensis Hodgson) – a predator of the Indian Giant Squirrel (Ratufa indica). Journal of the Bombay Natural History Society 91: 454.
Kannan, R. (1998). Avifauna of the Anaimalai Hills (Western Ghats) of southern India. Journal of the Bombay Natural History Society 95: 199.
Kimya, K., S. Satoshi & U. Teruaki (1989). Growth and development of the Black-eared Kite Milvus migrans lineatus. Japanese Journal of Ornithology 38: 31–42; http://doi.org/10.3838/jjo.38.31
Konig, C. & F. Weick (2010). Owls of the World. Christopher Helm Publishers, an imprint of A & C Black Publishers Ltd., London, 528pp.
Kumar, T.S. (1985). The Life History of the Spotted Owlet (Athene brama brama, Temminck) in Andhra Pradesh. Monograph of the Raptor Research Centre, Hyderabad, 241pp.
Llinas-Gutieerrez, J., G. Arnaud & M. Acevedo (1991). Food habits of the Great Horned Owl (Bubo virginianus) in the Cape Region of Lower California, Mexico. Journal of Raptor Research 25: 140–141.
Mani, M. (1990). General Entomology. Oxford & IBH Publishing, New Delhi, 912pp.
Mahmood-ul-Hassan, M., M.A. Beg & M. Mustaq-ul-Hassan (2007). Locality related changes in the diet of the Barn Owl (Tyto alba stertens) in agrosystems in Central Punjab, Pakistan. Wilson Journal of Ornithology 119: 479–483.
Marchesi, L. & P. Pedrini (2002). Biases associated with diet study methods in the Eurasian Eagle Owl. Journal of Raptor Research 36: 11–16.
Marshall, J.T. Jr. (1977). A synopsis of Asian species of Mus (Rodentia, Muridae). Bulletin of the American Museum of Natural History 158: 173–220.
Marshall, J.T. Jr. (1986). Systematics of the Genus Mus. Current Topics in Microbiology and Immunology 127: 12–18; http://doi.org/10.1007/978-3-642-71304-0_2
Martinez, J.A. (2003). Predictive models of habitat preference for the Eurasian Eagle Owl Bubo bubo: a multiscale approach. Ecography 26(1): 21–28.
Medelson, J.M. (1989). Habitat preferences, population size, food and breeding of six owl species in the Springbok Flats, South Africa. Ostrich 60(4): 183–190; http://doi.org/10.1080/00306525.1989.9633755
Mikkola, H. (1976). Owls killing and killed by other owls and raptors in Europe. British Birds 69: 144–154.
Mikkola, H. (2012). Owls of the World - A Photographic Guide. Christopher Helm, an imprint of Bloomsbury Publishing Plc, London, 512pp.
Mysterud, I. & H. Dunker (1982). Food and nesting ecology of the Eagle Owl Bubo bubo (L.) in four neighbouring territories in southern Norway. Swedish Sportsmen’s Ass. Swedish Wildlife Research (Viltrevy), 12pp.
Nel, J.A.J. (1969). The prey of owls of the Namib Desert, 1. The Spotted Eagle Owl Bubo africanus. Scientific Papers Namib Desert Research Station 43: 55–58.
Obuch, J. (2014). Spatial Diversity in the Diet of the Eurasian Eagle Owl Bubo bubo in Iran. Podoces 9: 7–21.
Orsini, P. (1985). Le regime alimentaire du Hibou grand-duc Bubo bubo en Provence. Alauda 53: 11–28.
Pande, S. & N. Dahanukar (2011). The diet of the Indian Eagle Owl Bubo bengalensis and its agronomic significance. Journal of Threatened Taxa 3(8): 2011–2017; http://doi.org/10.11609/JoTT.o2536.2011-7
Pande, S., A. Pawashe, M. Mahajan, A. Mahabal, C. Joglekar & R. Yosef (2011). Breeding biology, nesting habitat and diet of the Rock Eagle Owl (Bubo bengalensis). Journal of Raptor Research 45: 211–219; http://doi.org/10.3356/JRR-10-53.1
Penteriani, V., M. Gillardo & P. Roche (2002). Landscape structure and food supply affect Eagle Owl (Bubo bubo) density and breeding performance: a case of intra-population heterogeneity. Journal of Zoology 257: 365–372; http://doi.org/10.1017/S0952836902000961
Potapov, E. & R. Sale (2012). The Snowy Owl. T & AD Poyser, an imprint of Bloomsbury Publishing Plc., London, 304pp.
Prakash, I., P. Singh & A. Saravanan (1995). Ecological Distribution of Small Mammals in the Aravalli Ranges. Proceedings of the Indian National Scientific Academy 61: 137–148.
Ramanujam, M.E. (2004). Methods of analyzing rodent prey of the Indian Eagle Owl Bubo bengalensis (Franklin) in and around Pondicherry, India. Zoos’ Print Journal 19: 1492–1494; http://doi.org/10.11609/JoTT.ZPJ.1117a.1492-4
Ramanujam, M.E. (2006). On the prey of the Indian Eagle Owl Bubo bengalensis (Franklin, 1831) in and around Pondicherry, southern India. Zoos’ Print Journal 21: 2231–2240; http://doi.org/10.11609/JoTT.ZPJ.1425.2231-40
Ramanujam, M.E. (2015). Time budget and behavioural traits of adult and young Indian Eagle Owl Bubo bengalensis (Franklin, 1831) in and around a nesting site: a preliminary report. Journal of Threatened Taxa 7(14): 8139–8147+supplement; http://doi.org/10.11609/jott.2429.7.14.8139-8147
Ramanujam, M.E. & B. Verzhutskii (2004). On the prey of the Spotted Owlet Athene brama (Temminck) in a forested ravine in Auroville, Pondicherry. Zoos’ Print Journal 19(10): 1654–1655; http://doi.org/10.11609/JoTT.ZPJ.1093a.1654-5
Rasmussen, P.C. & J.C. Anderton (2012). Birds of South Asia – The Ripley Guide. Volume 2: Attributes and Status. Second Edition. National Museum of Natural History - Smithsonian Institution, Michigan State University and Lynx Edicions, Washington D.C., Michigan and Barcelona, 684pp.
Real, J. & S. Manosa (1990). Eagle Owl (Bubo bubo) predation on juvenile Bonelli’s Eagles (Hieraaetus fasciatus). Journal of Raptor Research 24: 69–71.
Robinson, M. & C.D. Becker (1986). Snowy Owls on Fetlar. British Birds 79: 228–242.
Rodel, H.G., W.W.A. Scholze & D. Kock (2002). Diet of Mackinder’s eagle owl Bubo capensis mackinderi in the alpine zone of Mount Kenya. African Journal of Ecology 40: 283–288.
Rosenberg, L.V. & R.J. Cooper (1990). Approaches to avian diet analysis. Avian Biology 13: 80–90.
Sergio, F. & A. Boto (1999). Nest dispersion, diet and breeding success of Black Kites (Milvus migrans) in the Italian alps. Journal of Raptor Research 33: 207–217.
Sergio, F. & P. Pedrini (2003). Adaptive selection of foraging and nesting habitat by black kites (Milvus migrans) and its implications for conservation: a multiscale approach. Biological Conservation 112: 351–362; http://doi.org/10.1016/S0006-3207(02)00332-4
Sergio, F., L. Marchesi & P. Pedrini (2003). Spatial refugia and the coexistence of a diurnal raptor with its intraguild owl predator. Journal of Animal Ecology 72: 232–245; http://doi.org/10.1046/j.1365-2656.2003.00693.x
Sergio, F. & F. Hiraldo (2008). Intraguild predation in raptor assemblages: a review. Ibis 150: 132–145; http://doi.org/10.1111/j.1474-919X.2008.00786.x
Seshadri, K.S. (2013). Lords of the Ravines - the Rock Eagle Owls of Pondicherry. Sanctuary Asia 33: 54–59.
Shehab, A.H. (2004). Diet of the Eagle Owl, Bubo bubo, in Syria. Zoology in the Middle East 33: 21 – 26.
Shehab, A.H. & M. Ciach (2008). Diet Composition of the Pharaoh Eagle Owl, Bubo ascalaphus, in Azraq Nature Reserve, Jordan. Turkish Journal of Zoology 32: 65–69.
Simeonov, S., B. Milchev & Z. Boev (1998). Study of the Eagle Owl (Bubo bubo L.) (Aves: Strigiformes) in the Strandzha Mountain (southeast Bulgaria). II. Food spectrum and trophic specialization. Acta Zoologica 50: 87–100.
Simmons, R.E., D.M. Avery & G. Avery (1991). Biases in diets determined from pellets and remains: correction factors for a mammal and bird-eating raptor. Journal of Raptor Research 26: 63–67.
Srinivasulu, C. & M.S. Pradhan (2003). Checklist of Murids (Mammalia: Rodentia: Muridae) of South Asia. Zoos’ Print Journal 18(12): 1286–1310; http://doi.org/10.11609/JoTT.ZPJ.18.12.1286-310
Srinivasulu, C., B. Srinivasulu, A. Rajesh, C.A.N. Rao & V. Nagulu (2004). Non-volant small mammals of Kasu Bramananda Reddy National Park, Andhra Pradesh. Zoos’ Print Journal 19: 1495–1497; http://doi.org/10.11609/JoTT.ZPJ.1088.1495-7
Steyn, P. & N. Myburgh (1983). Prey of the Cape Eagle Owl at Brandvlei, Cape. African Wildlife 37: 127.
Talmale, S.S. & M.S. Pradhan (2009). Identification of small Mammal species through Owl Pellet Analysis. Records of the Zoological Survey of India. Occasional Paper No. 294: 23pp+20pl.
Tella, J.L. & S. Manosa (1993). Eagle Owl predation on Egyptian Vulture and Northern Goshawk: possible effect of decrease in European Rabbit availability. Journal of Raptor Research 27: 111–112.
Trejo, A. & D. Grigera (1998). Food habits of the Great Horned Owl (Bubo virginianus) in a Patagonian Steppe in Argentina. Journal of Raptor Research 32: 306–311.
Tumurbat, J., G. Sundev & R. Yosef (2009). Nest site and food composition of the Eagle Owl Bubo bubo in Mongolia. Ardea 97: 419–523; http://doi.org/10.5253/078.097.0417
Vein, D. & M. Thevenot (1978). Etude sur le hibou Grand-Duc Bubo bubo ascalaphus dans le Moyen-Atlas Morocain. Nos Oiseaux 34: 347–351.
Verzhutskii, B. & M.E. Ramanujam (2002). On the prey of the Collared Scops Owl Otus bakkamoena (Pennant) at Auroville, Pondicherry. Zoos’ Print Journal 17(11): 939–940; http://doi.org/10.11609/JoTT.ZPJ.17.11.939-40
Viega, J.P. & F. Hiraldo (1990). Food habits and the survival and growth of nestlings in two sympatric kites (Milvus milvus and Milvus migrans). Ecography 13: 62–71.
Villafuerte, R. & J. Vinuela (2006). Size of Rabbits consumed by Black Kites increased after a Rabbit epizootic. Mammal Review 29: 261–264; http://doi.org/10.1046/j.1365-2907.1999.2940261.x