Physiological validation of enzyme immunoassay of fecal glucocorticoid metabolite levels and diurnal variation measured in captive Black-tufted Marmoset Callithrix penicillata (Mammalia: Primates: Callitrichidae)
Cristiane Schilbach Pizzutto 1, Manuela Gonçalves Fraga Geronymo Sgai 2, Cláudia Pereda Francischini 3, Priscila Viau 4, Cláudio Alvarenga de Oliveira 5 & Marcelo Alcindo de Barros Vaz Guimarães 6
1,2,3,4,5,6 Department of Animal Reproduction - Faculty of Veterinary Medicine and Zootechny - University of São Paulo - Brazil. Avenida Prof. Dr. Orlando Marques de Paiva, 87, Cidade Universitária, São Paulo, SP 055508-270, Brazil
1,2,6 Shape Brasil - Rua Doutor Miranda de Azevedo, 779 apto 104, Pompéia, São Paulo, SP; CEP: 05027-000, Brasil
1 cspizzutto@yahoo.com.br (corresponding author), 2 manuelafraga@hotmail.com, 3 capereda78@gmail.com, 4 priviau@usp.br, 5 cadolive@usp.br, 6 mabvg@usp.br
doi: http://dx.doi.org/10.11609/JoTT.o4099.7234-42
Editor: Ulrike Streicher, Wildlife Veterinarian / Wildlife Management Consultant, Eugene, USA Date of publication: 26 May 2015 (online & print)
Manuscript details: Ms # o4099 | Received 12 July 2014 | Final received 08 May 2015 | Finally accepted 10 May 2015
Citation: Pizzutto, C.S., M.G.F.G. Sgai, C.P. Francischini, P. Viau, C.A. de Oliveira & M.A. de Barros Vaz Guimarães (2015). Physiological validation of enzyme immunoassay of fecal glucocorticoid metabolite levels and diurnal variation measured in captive Black-tufted Marmoset Callithrix penicillata (Mammalia: Primates: Callitrichidae). Journal of Threatened Taxa 7(6): 7234–7242; http://dx.doi.org/10.11609/JoTT.o4099.7234-42
Copyright: © Pizzutto et al. 2015. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo) (10/51926-2 and 08/53562-8).
Competing interests: The authors declare no competing interests. Author Details: Cristiane Schilbach Pizzutto is veterinary post doctoral from University of São Paulo and works with animal welfare and environmental enrichment; she is coordinator of the Regional from The Shape of Enrichment (Shape Brasil). Manuela Gonçalves Fraga Geronymo Sgai is veterinary with doctoral from University of São Paulo; Claudia Pereda Francschini is biomedical with doctoral from Chemistry Institute from University of São Paulo; Priscila Viau is veterinary and she is technical from hormonal dosage laboratory at veterinary medicine school from University of São Paulo; Claudio Alvarenga de Oliveira is veterinary, professor and coordinator from hormonal dosage laboratory at Department of Animal Reproduction - Faculty of Veterinary Medicine and Zootechny - University of São Paulo; Marcelo Alcindo de Barros Vaz Guimarães is veterinary and professor of wild animals reproduction at Department of Animal Reproduction - Faculty of Veterinary Medicine and Zootechny - University of São Paulo Author Contribution: CSP and MABVG designed the project; CSP and MGFGS worked during all project at the collect of samples and dosage of hormones; CPF, PV and CAO helped at the hormonal dosage; CSP and MABVG analysed the datas; CSP wrote the article and MABVG was responsible for review the article and supervisor the project.
Acknowledgements: The authors would like to thank FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo) for the financial support (10/51926-2 and 08/53562-8).
Abstract: Measuring stress responses is an important aspect for the conservation of endangered wild species. Non-invasive measuring of glucocorticoid metabolite levels has become an important tool to measure stress intensity. The aims of the present study were as follows: to validate the enzyme immunoassay to measure the concentration of fecal metabolites of glucocorticoids (FGM) after stressful stimuli and to determine whether FGM concentrations fluctuate diurnally in Black-tufted Marmosets Callithrix penicillata in captivity. Eight captive healthy adult Black-tufted Marmosets (four males and four females) were included in the study. The animals were subjected to three treatments: (1) hormone challenge with adrenocorticotropic hormone (ACTH), (2) saline administration and (3) control treatment to monitor diurnal changes of FGM. Fecal samples were collected on days -1, 0, +1 and +2, with intramuscular administration of ACTH and saline performed on day 0. To control diurnal variations, all feces from all animals were collected over six consecutive days and identified using the time of defecation and animal identification number. There were four designated two-hour periods per day (8–10 h, 10–12 h, 12–14 h and 14–16 h), and the samples were grouped for each two-hour period to obtain a representative pool. The samples were frozen, and the metabolite concentrations were measured by enzyme immunoassay following extraction. The results show that immunoassay measurements of FGM concentrations in C. penicillata can be validated physiologically. Diurnal variation of the FGM concentration was observed, with significantly increased FGM levels in the early afternoon in both sexes. The mean FGM concentration was higher in captive females than in males. Physical restraint followed by saline administration led to adrenocortical stimulation similar to that observed following ACTH hormone challenge, a finding that has not previously been reported in C. penicillata. Our results show immunoassay measurements of FGM concentrations provide a valuable tool for the non-invasive study of the endocrine correlates of behavior and well-being of this species.
Keywords: Callithrix penicillata, diurnal variation, fecal glucocorticoid metabolites, stress, validation.
Portuguese Abstract: As mensurações das respostas ao estresse são aspectos importantes para a conservação de espécies selvagens ameaçadas de extinção. O monitoramento não invasivo tornou-se uma ferramenta importante para a mensuração de metabólitos de glicocorticoides, como forma indireta de avaliação da intensidade do estresse. Os objetivos deste estudo foram: validar o enzima imunoensaio para a mensuração das concentrações de metabólitos fecais de glicocorticoides (MFG), após a realização de estímulos estressantes e determinar as variações diurnas das concentrações destes metabólitos, em sagui-de-tufo-preto (Callithrix penicillata), mantidos em cativeiro. Oito saudáveis adultos cativos de sagui-de-tufo-preto (4 machos e 4 fêmeas) foram incluídos neste estudo. Os animais foram submetidos a 3 tratamentos a saber: (1) Desafio hormonal com aplicação de ACTH, (2) Aplicação de solução salina e (3) Controle para acompanhamento das variações diurnas. Amostras fecais foram coletadas nos dias -1, 0, +1 e +2, sendo o dia 0, o dia das aplicações intramusculares tanto de ACTH quanto de solução salina. Para o controle da variação diurna, todas as fezes dos animais foram coletadas por seis dias consecutivos e identificadas usando a hora de defecação e o número de identificação do animal. O delineamento foi feito em quatro períodos de duas horas por dia (8-10h, 10-12h, 12-14h and 14-16h), e as amostras de cada período de duas horas foram agrupadas obtendo-se uma média. As amostras foram congeladas e depois da extração, mensuradas as concentrações de metabólitos por enzima imunoensaio. Os resultados mostraram que foi possível validar fisiologicamente o imunoensaio para a mensuração de MFG em C. penicillata,. Foi caracterizada a existência de variação diurna, com elevação significativa de suas concentrações no início da tarde, em ambos os sexos. A contenção física seguida da administração de solução salina promoveu estimulação adrenocortical semelhante à obtida no desafio hormonal com ACTH. Fêmeas cativas de C. penicillata apresentam maiores concentrações médias de MFG do que machos. Os resultados descritos oferecem uma valiosa ferramenta para o estudo não-invasivo da endocrinologia comportamental e do bem estar da espécie.

Introduction
Many studies in the past, have focussed on understanding the implications of stress on vertebrate species better. As described by Heistermann et al. (2006), chronic stress may have deleterious effects on physiological, psychosocial and behavioral parameters. Chronic stress may also compromise immune functions by inhibiting the release of cytokines and lymphocytes and consequently increasing disease susceptibility (Sapolsky 2002; Wingfield 2005). Other studies have reported suppressed reproductive functions (Wingfield & Sapolsky 2003) and effects on cognitive abilities (Ohl & Fuchs 1999) and behavior (Wielebnowski et al. 2002) resulting from chronic stress. The combination of animal behavior analysis and the measurement of glucocorticoid levels is a promising approach (Pizzutto et al. 2010) to study the impacts of stress (Romero 2004).
Glucocorticoids are steroid hormones produced by the adrenal glands, and the major glucocorticoids are cortisol and corticosterone. Following the immediate adrenergic response that releases catecholamines in situations of acute stress, glucocorticoids increase the amount of energy available to the body for necessary physiological functions, such as locomotion (Sapolsky 2002; Reeder & Kramer 2005).
In addition to mediating the stress response, glucocorticoids are essential to regulate energy availability based on activity patterns. For example, diurnal animals with highly circadian activity patterns have increased glucocorticoid levels at the end of the dark cycle, which provides energy for locomotion and foraging activities. In nocturnal animals, the rise in glucocorticoid levels occurs at the end of the light cycle (Reeder & Kramer 2005; Touma & Palme 2005). It is, therefore, critical to take into account the circadian rhythms of a species when standardizing the time of sample collection for glucocorticoid measurements (Heintz et al. 2011). In this vein, the application and validation of non-invasive hormone measurement techniques reflect the need to complement behavioral data with endocrine-physiological information (Heintz et al. 2011).
Glucocorticoids and their metabolites can be reliably measured in the blood, urine and feces of many species using immunoenzymatic assays (Romano et al. 2010; Sheriff et al. 2011). For the study of stress physiology in wild animals, non-invasive methods based on the measurement of glucocorticoid metabolites in urine or feces have become the preferred approach because many samples can be collected over long periods of time in a safe and non-stressful manner. The physical restraints required for the serial collection of blood samples are undesirable because they induce stress and are sometimes difficult to perform due to the physical and behavioral characteristics of a given species (Heistermann et al. 2006; Rimbach et al. 2013). When analyzing changes in the concentrations of glucocorticoid metabolites, particularly in the feces, the pulsatile changes that occur in the blood levels of these metabolites over a period of hours or days have to be taken into account by frequently measuring metabolite concentrations (Whitten et al. 1998; Rimbach et al. 2013). These non-invasive methods have been widely used in reproductive (Strier & Ziegler 1994, 1997; Brown et al. 1995; Schwarzenberger et al. 1996; Ziegler et al. 1997), behavioral and welfare studies (Pizzutto et al. 2008a,b, 2010).
Despite the tremendous progress in the application of non-invasive monitoring techniques, several problems related to urinary or fecal metabolite measurements remain unresolved (Higham et al. 2010). For example, the considerable differences in steroid metabolism in different species make it quite difficult to determine the prevalent metabolite (fecal or urinary) and the appropriate antibody for measurement assays (Palme et al. 2005). Therefore, validation in different species is critical for the different methods to measure glucocorticoid metabolites (Palme et al. 2005; Touma & Palme 2005), particularly for techniques involving the application of exogenous adrenocorticotropic hormone (ACTH).
There is a lack of information in literature regarding the physiology of the Black-tufted Marmoset Callithrix penicillata (Image 1) a neotropical primate species with a great potential as a model for reproductive and behavioral studies. The aims of the present study were as follows: (1) to validate the assay used to measure fecal glucocorticoid metabolites (FGM) in Black-tufted Marmosets and (2) to determine whether the concentrations of these metabolites fluctuate diurnally.

Methods
Animals
Eight adult Black-tufted Marmosets (C. penicillata) (four males and four females) were included in the study. All had intact reproductive systems and had been kept in the Wild Animal Rehabilitation Center (Centro de Reabilitação de Animais Silvestres, CRAS) of the Department of Parks and Green Areas (Departamento de Parques e Áreas Verdes, DEPAVE III) of the municipality of São Paulo, São Paulo, Brazil. The animals were separated into pairs and kept in 120cm x 60cm x 60cm cages. While all of the animals had auditory, visual and olfactory contact with each other, physical contact was restricted to their respective partners. Following the standard protocols of the supporting institution food was provided twice a day, in the early morning and late afternoon. The diet consisted primarily of seasonal fruits, primate feed (MegaZoo®, Minas Gerais, MG, Brazil), meal worms, crickets and eggs. Water was available ad libitum. All of the animals had been in captivity for at least two years by the time the study was performed.
Physiological validation
All of the animals were subjected consecutively to the three following treatments: Treatment 1—ACTH: hormone challenge with ACTH administration; Treatment 2—saline: saline administration; and Treatment 3—control: no intervention. Treatment 3 was used to monitor diurnal changes.
Treatment 1—hormone challenge with ACTH administration: Over four consecutive days, all feces from all animals were collected and labelled with the time of defecation and animal identification number. With day 0 corresponding to the day of ACTH administration, the collection of fecal samples began on day-1 and continued on days 0, +1 and +2. As it was unpredictable to determine when each individual would defecate, there were four designated two-hour periods per day (8–10 h, 10–12 h, 12–14 h and 14–16 h), and the samples were grouped for each two-hour period from each animal to obtain a representative pool. On day 0 at 8–10 h, the animals were physically restrained for intramuscular (IM) administration of 0.3 ml Synacthen Depot® of a 0.1 % solution (0.1g/100ml), which corresponds to 31 IU/kg ACTH (Heistermann et al. 2006).
Treatment 2—saline administration: Over four consecutive days, all feces from all animals were collected and labelled with the time of defecation and animal identification number. Collection began on day-1 and continued on days 0, +1 and +2. There were four designated two-hour periods per day (8–10 h, 10–12 h, 12–14 h and 14–16 h), and the samples from each animal were grouped for each two-hour period to obtain a representative pool. On day 0 at 8–10 h, the animals were physically restrained for intramuscular administration of 0.3ml of a saline solution 0.9%.
Treatment 3—control to monitor diurnal changes: Over six consecutive days, all feces from all animals were collected and labelled with the time of defecation and animal identification number. There were four designated two-hour periods per day (8–10 h, 10–12 h, 12–14 h and 14–16 h), and the samples were grouped for each two-hour period to obtain a representative pool. During these six days, the animals were not restrained, and no substances were administered to them. To avoid interference between consecutive treatments and to provide a resting period for the animals, the treatments were separated by one-week intervals.
A 2ml dose of Arcolor® food coloring for chocolate was mixed with fruit juice orally administered twice a day to all females to differentiate their feces from that of the males in the study. Samples were collected from 8–16 h, at the end of the designated two hour periods; after this the cages were placed in a closed space to allow the animals to sleep. Feces were not found during night-time, and the first morning samples were produced shortly after the animals awoke.
All samples were stored in a freezer at -200C until they were processed at the Hormone Dosage Laboratory (Laboratório de Dosagens Hormonais, LDH) of the Department of Animal Reproduction (Departamento de Reprodução Animal, DRA) of the School of Veterinary Medicine and Animal Husbandry (Faculdade de Medicina Veterinária e Zootecnia, FMVZ), University of São Paulo (Universidade de São Paulo, USP).
Fecal extraction and sample processing
The metabolites were extracted according to the technique described by Ziegler et al. (1996) with some modifications. For each sample, 0.2g of the lyophilized (freeze-dried) feces were weighed and transferred to an appropriately labeled test tube. A 5ml volume of an 80% ethanol (MERCK®) solution prepared with type I water (Milli-Q®) was added. The tubes were placed in a multivortex device (VWR Scientific Products, VX-2500) for 15 minutes for homogenization, followed by centrifugation (CENTRIBIO, TDL80-2B) for 15 minutes at 3000 rpm. A 1.5ml volume of supernatant (fecal extract) was transferred to a 2.0ml plastic tube and stored in a freezer at -300C.
Prior to the analysis, the samples were diluted in gelatin buffer (pH 7.0) (13.8g NaPO4, 9.0g NaCl, 1.0g sodium azide and 1.0 g gelatin in 1000ml distilled water).
Hormone analyses
The samples were analyzed using the parallelism test. Twenty samples with high concentrations were diluted to obtain concentration values similar to those used to develop the calibration curve for the assay. For this purpose fecal samples were serially diluted in the same proportions as the calibration curves for the assay. Depleted fecal extract steroids were used as the matrix. The curve obtained was compared to the calibration curve using a linear regression to assess parallelism of the two curves.
FGM concentrations were measured using an enzyme immunoassay method (EIM) following procedures described for other mammals (Heistermann et al. 2006), using cortisol R4866 monoclonal antibodies (University of California, Davis, USA) as described by Munro & Stabenfeldt (1984).
Statistical analysis
The data were analyzed to assess whether the residues exhibited normal distributions, and a homogeneity of variance test was performed using the Guided Data Analysis application with SAS/LAB® Guided Data Analysis application.
The mean glucocorticoid concentration was determined using all samples of each individual in each group. All samples with values two standard deviations (SD) above the mean were removed. The mean was recalculated, and this process was repeated until all values were within two SD from the mean. The resulting mean value was designated the baseline value, and all values greater than two SD from the mean were considered peak values. This process allows for a statistical assessment of baseline values for biologically relevant hormone concentrations (Heintz et al. 2011).
For each period of diurnal variation studied, the mean FGM concentration and standard deviation were calculated. The means were compared using ANOVA and Tukey’s test, both with P<0.05.
The present study was approved by the Bioethics Commission of the School of Veterinary Medicine and Animal Science, São Paulo University (FMVZ, USP), Brazil under procedure number 1470/2008.
Results
The values obtained for the FGM parallelism test were Y = 5.485+0.93 * X and R2 = 0.984 showing that the antibody used in the assays was highly capable to detect the searched metabolite in the fecal matrix.
For the hormone tests, the low and high inter-assay variation values were 5.5% and 10.1%, respectively, and the low and high intra-assay variation coefficients were 2.3% and 5.6%, respectively. The sensitivity of the assay was 2.1 ng/well, measured in the calibration curve.
Treatment 1 (hormone challenge with ACTH administration)
The mean basal FGM concentrations and standard deviations for the females were 636.41 (±314.58) and for the males 255.20 (±113.50) ng/g dry feces. Thus, prior to ACTH administration, females had mean FGM levels 2.49 times higher than males.
After ACTH administration, the FGM levels reached the peak values after approximately 9.25 hours in females and 8.75 hours in males (Fig. 1). The peak FGM values were 2.47 and 2.20 times greater than the basal values in females and males, and the FGM concentrations returned to basal levels after 32 hours in both sexes.
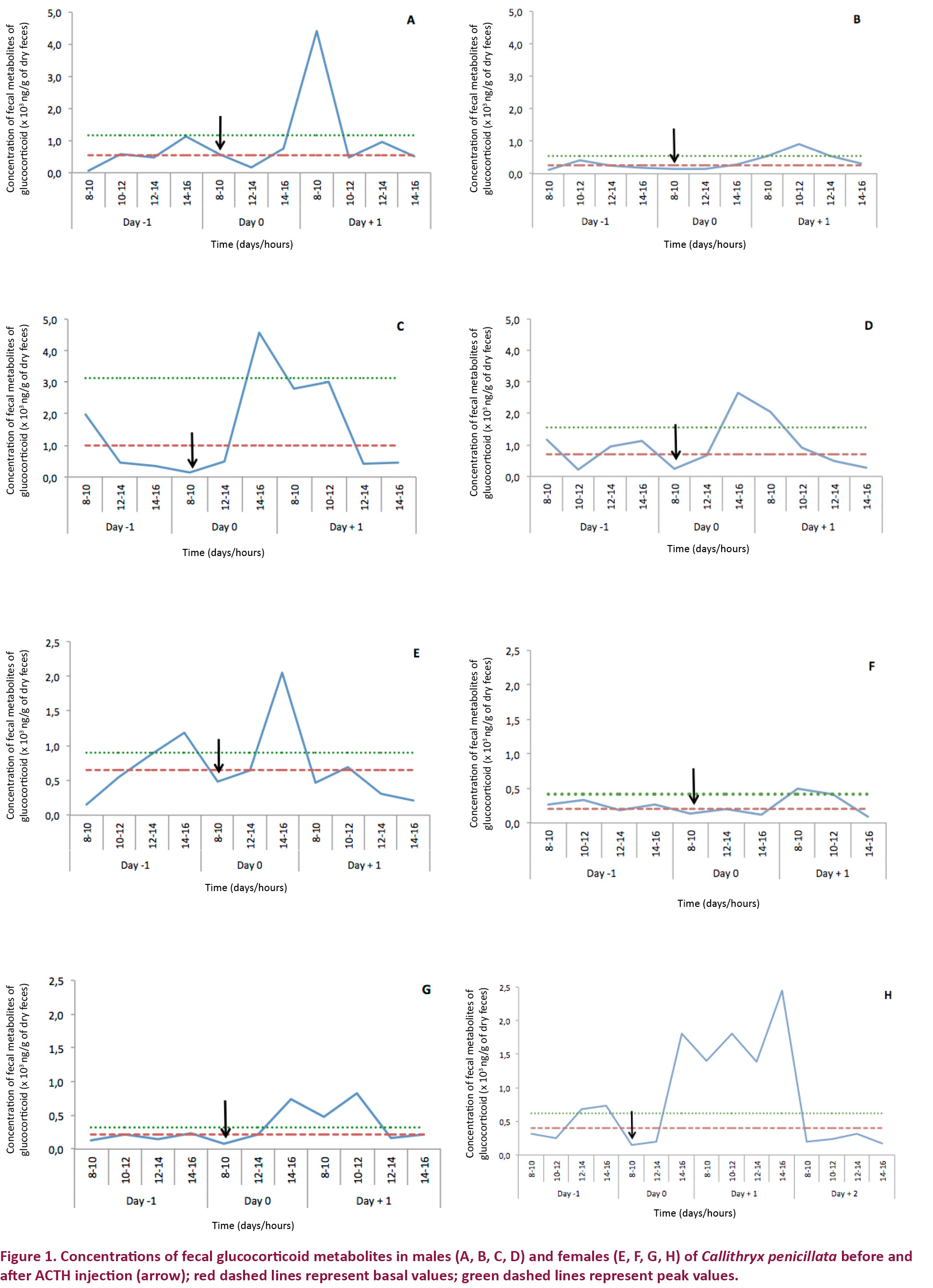

Treatment 2 (saline administration)
The mean basal FGM concentrations in females were 506.18 (±470.12) and in males 278.95 (±89.42) ng/g dried feces, indicating that the FGM values were 1.81 times greater in females than in males prior to saline administration.
Following saline administration, the FGM levels exceeded the basal values after six hours in both sexes (Fig. 2), and the peak values were observed after approximately 9.25 hours in females and 13.50 hours in males. The peak FGM values were 1.70 times higher than the basal values in females and 1.92 times higher in males. The FGM concentrations returned to basal levels after 34.5 hours in females and after 28 hours in males.
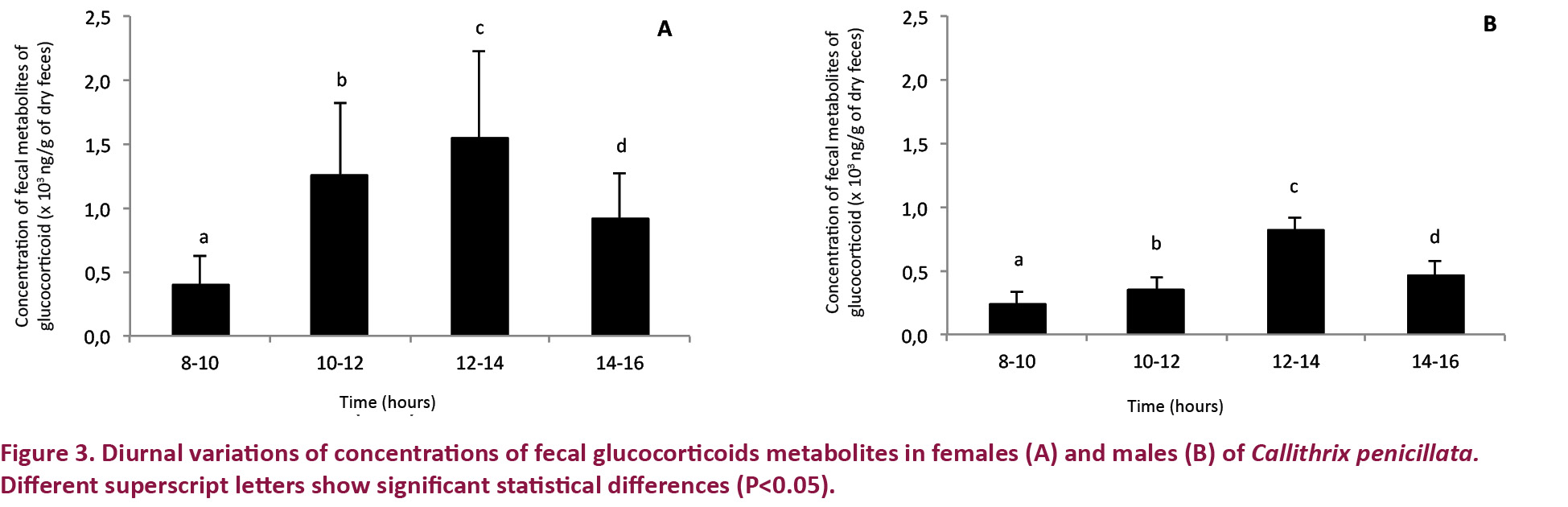
Treatment 3 (control for monitoring diurnal changes)
In terms of diurnal fluctuation, the FGM concentrations in both females and males were highest in the afternoon (12–14 h) during the study periods (Fig. 3). In the afternoons, the mean FGM concentrations were 1.89 times greater in females than in males.
Discussion
This study represents the first to validation of immunoassays for the measurement of FGM concentrations in Black-tufted Marmosets (C. penicillata).
It was previously reported that cortisol metabolites in C. jacchus are primarily excreted in the urine (Bahr et al. 2000). The results described here, however, clearly show an increase in FGM levels after the administration of either ACTH or saline solution. The diurnal fluctuation observed was consistent with the findings of Sousa & Ziegler (1998), who emphasized the importance to start to collect samples shortly after the animals wake up in order to monitor diurnal changes in the adrenal response. Considering the criteria described by Wasser et al. (2000), Touma & Palme (2005) and Heistermann et al. (2006), the results of the present study demonstrate that FGM measurements in C. penicillata can be physiologically validated and that changes in adrenocortical activity can be safely monitored with this method.
Although little is known about the diurnal variation of plasma cortisol in marmosets, Coe & Levine (1995) observed fluctuations in cortisol concentrations in Saimiri sciureus, that were very similar to those reported in other diurnal primates, namely, maximum cortisol concentrations in the morning and minimum concentrations in the late afternoon. The results of our study, however, were consistent with the findings of Souza & Ziegler (1998) in C. jacchus and we observed higher FGM concentrations in the early afternoon (12–14 h) and significantly lower values in the morning (8–10 h) in all animals.
The difference in time between the morning increase in plasma cortisol described in the literature and the afternoon increase in FGM concentration observed in the present study can be attributed to the amount of time required for the metabolization of the glucocorticoids in the blood and their fecal excretion and the resulting time interval before the glucocorticoids are converted into fecal metabolites (Ziegler et al. 2009). Thus, morning plasma increases are manifested as FGM increases in the early afternoon. This time delay is known to be affected by diet composition, liver function and gastrointestinal passage (Goldin et al. 1982; Wasser et al. 1993). According to Touma & Palme (2005) and Rimbach et al. (2013), diurnal FGM changes may be impossible to evaluate in animals with excessively long intestines and long intervals between defecation episodes. In the present study, the profile of changes in FGM concentration throughout the day were similar in males and females (Fig. 3).
Similar adrenocortical responses were observed for the ACTH and saline administrations, indicating that both treatments caused stress in some way. Moberg (1985) and Smith & French (1997) described how physical restraint of animals followed by an intramuscular injection can be considered a stressor capable of inducing increased glucocorticoid production. This view is clearly supported by our results, given that the FGM increase following saline administration was similar to that observed following the ACTH challenge. The time intervals observed for peak FGM concentrations were consistent with the range of 8–24 hours reported by Bahr et al. (2000) for C. jacchus.
Another important finding of the present study were the consistently higher FGM concentrations in females than in males. Possible explanations for this observation could be differences in the metabolic rate between males and females as discussed by Palme et al. (2005), Touma et al. (2003) and Belz et al. (2003). We also hypothesize that these results may reflect the relatively small sample size of the study or, considering the high dependence of the physiological impact of stressors on the perception of environmental stimuli, a greater difficulty to adapt to captivity among the females studied (Natelson et al. 1987).
In conclusion, the enzyme immunoassay used is a reliable method to measure FGM in captive C. penicillata.
REFERENCES
Bahr, N.I., R, Palme, U, Möhle, J.K. Hodges & M. Heistermann (2000). Comparative aspects of the metabolism and excretion of cortisol in three individual nonhuman primates. General and Comparative Endocrinology 117: 427–438; http://dx.doi.org/10.1006/gcen.1999.7431
Belz, E.E., J.S. Kennell, R.K. Czambel, R.T. Rubin & M.E. Rhodes (2003). Environmental enrichment lowers stress-responsive hormones in singly housed male and female rats. Pharmacology, Biochemistry and Behavior 76: 481–486; http://dx.doi.org/10.1016/j.pbb.2003.09.005
Brown, J.L., C.M. Wemmer & J. Lehnhardt. (1995). Urinary cortisol analysis for monitoring adrenal activity in elephants. Zoo Biology 14: 533–542; http://dx.doi.org/10.1002/zoo.1430140606
Coe, C.L. & S. Levine (1995). Diurnal and annual variation of adrenocortical activity in the squirrel monkey. American Journal of Primatology 35: 283–292; http://dx.doi.org/10.1002/ajp.1350350404
Goldin, B.R., H. Adlercreutz, S.L. Gorbach, J.H. Warram, J.T. Dwyer, L. Swenson & M. Woods (1982). Estrogen excretion patterns and plasma levels in vegetarian and omnivorous women. The New England Journal of Medicine 307(25): 1542–1547.
Heintz, M.R., R.M. Santymire, L.A. Parr & E.V. Lonsdorf (2011). Validation of a Cortisol Enzyme Immunoassay and Characterization of Salivary Cortisol Circadian Rhythm in Chimpanzees (Pan troglodytes). American Journal of Primatology 73: 903-908; http://dx.doi.org/10.1002/ajp.20960
Heistermann, M., R. Palme & A. Ganswindt (2006). Comparison of different enzymeimmunoassays for assessment of adrenocortical activity in primates based on fecal analysis. American Journal of Primatology 68: 257–273; http://dx.doi.org/10.1002/ajp.20222
Higham, J.P., A.B. Vitale, A.M. Rivera, J.E. Ayala & D. Maestripieri (2010). Measuring salivary analytes from free-ranging monkeys. Physiology and Behavior 101: 601–607; http://dx.doi.org/10.1016/j.physbeh.2010.09.003
Moberg, G.P. (1985). Animal Stress. American Physiological Society: Distributed by Williams & Wilkins, Bethesda, Md., Baltimore, viii+324pp.
Munro, C. & G. Stabenfeldt (1984). Development of microtitre plate enzyme-immunoassay for the determination of progesterone. Journal of Endocrinology 101: 41–49.
Natelson, B.H., D. Creigghton, R. MC Carty, W.N. Tapp, D. Pitman & J.E. Ottenweiler (1987). Adrenal hormonal indices of stress in laboratory rats. Physiology and Behavior 39: 117–125; http://dx.doi.org/10.1016/0031-9384(87)90408-2
Ohl, F. & E. Fuchs (1999). Differential effects of chronic stress an memory processes in the three shrew. Cognitive Brain Research 7: 379–387; http://dx.doi.org/10.1016/S0926-6410(98)00042-1
Palme, R., S. Rettenbacher, C. Touma, S.M. El-Bahr & E. Mostl (2005). Stress hormones in mammals and birds: Comparative aspects regarding metabolism, excretion and non- invasive measurement in fecal samples. Annals New York Academy of Sciences 1040: 162–171.
Pizzutto, C.S., M.G.F.G. Sgai, P. Viau, M.O.M. Chelini, C.A. Oliveira & M.A.B.V. Guimarães (2008a). Validação laboratorial e fisiológica de conjunto comercial para a quantificação de corticoides fecais em chimpanzé (Pan troglodytes) e orangotango (Pongo pygmaeus), cativos e submetidos a enriquecimentos ambientais. Brazilian Journal of Veterinary Research and Animal Science 45: 104–110; http://dx.doi.org/10.1590/S1413-95962008000700015
Pizzutto, C.S., M. Nichi, M.G.F.G. Sgai, S.H.R. Correa, P. Viau, A.M. Beresca, C.A. Oliveira, R.C. Barnabe & M.A.B.V. Guimarães (2008b). Effect of environmental enrichment on behavioral and endocrine aspects of captive Orangutan (Pongo pygmaeus). Laboratory Primate Newsletter 47: 10–14.
Pizzutto, C.S., M.G.F.G. Sgai, S.H.R. Correa, A.M. Beresca, P.V. Furtado, C.A. Oliveira, M. Nichi & M.A.B.V. Guimarães (2010). Enriquecimento ambiental e condicionamento operante com reforço positivo no retorno da ciclicidade ovariana de uma fêmea de chimpanzé (Pan troglodytes) - relato de caso. Clínica Veterinária 85: 66–72.
Reeder, D.M. & K.M. Kramer (2005). Stress in free-ranging mammals: integrating physiology, ecology, and natural history. Journal of Mammalogy 86: 225–235; http://dx.doi.org/10.1644/BHE-003.1
Rimbach, R., E.W. Heymann, A. Link & M. Heistermann (2013). Validation of an enzyme immunoassay for assessing adrenocortical activity and evaluation of factors that affect levels of fecal glucocorticoid metabolites in two New World primates. General and Comparative Endocrinology 191: 13–23; http://dx.doi.org/10.1016/j.ygcen.2013.05.010
Romano, M.C., A.Z. Rodas, R.A. Valdez, S.E. Hernandez, F. Galindo, D. Canales & D.M. Brousset (2010). Stress in wildlife species: noninvasive monitoring of glucocorticoids. Neuroimmunomodulation 17: 209–212; http://dx.doi.org/10.1159/000258726
Romero, L.M. (2004). Physiological stress in ecology: lessons from biomedical research. Trends in Ecology & Evolution 19: 249–255; http://dx.doi.org/10.1016/j.tree.2004.03.008
Sapolsky, R.M. (2002). Endocrinology of the stress-response, pp. 409–450. In: Becker, J.B., S.M. Breedlove, D. Crews, M.M. McCarthy (eds.). Behavioral Endocrinology. MIT Press, Cambridge, xxvi+806pp.
Schwarzenberger, F., E. Mostl, R. Palme & E. Bamberg (1996). Faecal steroid analysis for non-invasive monitoring of reproductive status in farm, wild and zoo animals. Animal Reproduction Science 42: 515–526; http://dx.doi.org/10.1016/0378-4320(96)01561-8
Sheriff, M.J., B. Dantzer, B. Delehanty, R. Palme & R. Boonstra (2011). Measuring stress in wildlife: techniques for quantifying glucocorticoids in wildlife. Oecologia 166: 869–887; http://dx.doi.org/10.1007/s00442-011-1943-y
Smith, T.E. & J.A. French (1997). Psychosocial stress and urinary cortisol excretion in Marmoset Monkeys (Callithrix kuhli). Physiology and Behavior 62: 225–232; http://dx.doi.org/10.1016/S0031-9384(97)00103-0
Sousa, M.B.C. & T.E. Ziegler (1998). Diurnal variation on the excretion patters of fecal steroids in Common Marmoset (Callithrix jacchus) females. American Journal of Primatology 46: 105–117; http://dx.doi.org/10.1002/(SICI)1098-2345(1998)46:2<105::AID-AJP1>3.0.CO;2-#
Strier, K.B. & T.E. Ziegler (1994). Insights into ovarian function in wild Muriqui Monkeys (Brachyteles arachnoides). American Journal of Primatology 32: 31–40; http://dx.doi.org/10.1002/ajp.1350320104
Strier, K.B. & T.E. Ziegler (1997). Behavioral and endocrine characteristics of the reproductive cycle in wild Muriqui Monkeys, Brachyteles arachnoids. American Journal of Primatology 42: 299–310; http://dx.doi.org/10.1002/(SICI)1098-2345(1997)42:4<299::AID-AJP5>3.0.CO;2-S
Touma C., N. Sachser, E. Möstl & R. Palme (2003). Effects of sex and time of day on metabolism and excretion of corticosterone in urine and feces of mice. General and Comparative Endocrinology 130: 267–278; http://dx.doi.org/10.1016/S0016-6480(02)00620-2
Touma C. & R. Palme (2005). Measuring fecal glucocorticoid metabolites in mammals and birds: the importance of validation, pp. 54–74. In: Bauchinger U., W. Goymann & S.J. Eiermann (eds.). Bird Hormones and Bird Migrations: Analyzing Hormones in Droppings and Egg Yolks and Assessing Adaptations in Long-distance migration. Annals of the New York Academy of Sciences (1046), 246pp.
Wasser, S.K., R. Thomas, P.P. Nair, C. Guidry, J. Southers, J. Lucas, D.E. Wildt & S.L. Monfort (1993). Effects of dietary fibre on faecal steroid measurements in Baboons (Papio cynocephalus). Journal of Reproduction and Fertility 97(2): 569–574.
Wasser, S.K., K.E. Hunt, J.L. Brown, K. Cooper, C.M. Crockett, U. Bechert, J.J. Millspaugh, S. Larson & S. Monfort (2000). A generalized fecal glucocorticoid assay for use in a diverse array of nondomestic mammalian and avian species. General and Comparative Endocrinology 120(3): 260–275; http://dx.doi.org/10.1006/gcen.2000.7557
Whitten, P.L., R. Stavisky, F. AurelI & E. Russell (1998). Response of fecal cortisol to stress in captive Chimpanzees (Pan troglodytes). American Journal of Primatology 44: 57–69; http://dx.doi.org/10.1002/(SICI)1098-2345(1998)44:1<57::AID-AJP5>3.0.CO;2-W
Wielebnowski, N.C., K. Ziegler, D.E. Wildt, J. Lukas & J.L. Brown (2002). Impact of social management on reproductive, adrenal and behavioural activity in the Cheetah (Acinonyx jubatus). Animal Conservation 5: 291–301; http://dx.doi.org/10.1017/S1367943002004043
Wingfield, J.C. (2005). The concept of allostasis: coping with a capricious environment. Journal of Mammalogy 86: 248–254; http://dx.doi.org/10.1644/BHE-004.1
Wingfield, J.C. & R.M. Sapolsky (2003). Reproduction and resistance to stress: when and how. Journal of Neuroendocrinology 15: 711–724; http://dx.doi.org/10.1046/j.1365-2826.2003.01033.x
Ziegler, T.E., C.T. Snowdon, D.H. Abbott, G. Scheffer, D.J. Wittwer & N.J. Schultz-Darken (1996). The metabolism of reproductive steroids during the ovarian cycle in two species of Callithrichids, Saguinus oedipus and Callithrix jacchus and estimation of the ovulatory period from fecal steroids. Biology of Reproduction 54: 91–99.
Ziegler, T.E., C.V. Santos, A. Pissinati & K.B. Strier (1997). Steroid excretion during the ovarian cycle in captive and wild Muriqui (Brachyteles arachnoids). American Journal of Primatology 42: 311-321; http://dx.doi.org/10.1002/(SICI)1098-2345(1997)42:4<311::AID-AJP6>3.0.CO;2-#
Ziegler, T.E., K.B. Strier & S. van Belle (2009). The reproductive ecology of South American primates: eco- logical adaptations in ovulation and conception, pp. 191–210. In: Garber, P.A., A. Estrada, J.C. Bicca-Marques, E.W. Heymann & K.B. Strier, (eds.). South American Primates. Comparative Perspectives in the Study of Behavior, Ecology, and Conservation, New York, Springer, New York, xvi+564pp.