Pseudophilautus dilmah, a new species of shrub frog (Amphibia: Anura: Rhacophoridae) from a threatened habitat Loolkandura in Sri Lanka
L.J. Mendis Wickramasinghe 1, Imesh Nuwan Bandara 2, Dulan Ranga Vidanapathirana 3, Kamani H. Tennakoon 4, Sameera R. Samarakoon 5 & Nethu Wickramasinghe 6
1,2,3,6 Herpetological Foundation of Sri Lanka, 31/5, Alwis Town, Hendala, Wattala, Sri Lanka
2 Ellangàwa Unity Care for Community and Nature, Hapugoda, Ambathenna, Sri Lanka
4,5,6 Institute of Biochemistry, Molecular Biology, Biotechnology, University of Colombo, Sri Lanka
1 boiga2000@gmail.com (corresponding author), 2 imeshnu1@gmail.com, 3 dulanrangavp@gmail.com,
4 kamani@ibmbb.cmb.ac.lk, 5 ranganath148@yahoo.com, 6 nemzy821@gmail.com
doi: http://dx.doi.org/10.11609/JoTT.o3501.7089-110 | ZooBank: urn:lsid:zoobank.org:pub:129EB17A-2F4E-42C9-995C-10E012E337D1
Editor: Annemarie Ohler, Muséum national d’Histoire naturelle, Paris, France. Date of publication: 26 April 2015 (online & print)
Manuscript details: Ms # o3501 | Received 27 January 2013 | Final received 12 March 2015 | Finally accepted 10 April 2015
Citation: Wickramasinghe, L.J.M., I.N. Bandara, D.R. Vidanapathirana, K.H. Tennakoon, S.R. Samarakoon & N. Wickramasinghe (2015). Pseudophilautus dilmah, a new species of shrub frog (Amphibia: Anura: Rhacophoridae) from a threatened habitat Loolkandura in Sri Lanka. Journal of Threatened Taxa 7(5): 7089–7110; http://dx.doi.org/10.11609/JoTT.o3501.7089-110
Copyright: © Wickramasinghe et al. 2015. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: Biodiversity Secretariat of the Ministry of Environment, Nagao Natural Environment Foundation, and Dilmah Conservation.
Competing Interest: The authors declare no competing interests.
Author Details: L.J. Mendis Wickramasinghe, is the founder, President of the Herpetological Foundation of Sri Lanka, and is the Principal Investigator in the current project. He has close to two decades of field herpetological (reptile and amphibian) experience in Sri Lanka with a focus on taxonomic identifications. Imesh Nuwan Bandara, M.Sc, B.Sc specializing in Zoology at the University of Peradeniya, Sri Lanka, with a keen interest in sustainable development, community based conservation, traditional agricultural practices, ethnobotany, local biodiversity and behavioral ecology of herpetofauna and other wild fauna. Dulan Ranga Vidanapathirana, is the Vice-President, and a founder member of the Herpetological Foundation of Sri Lanka, with over 10 years of field herpetological, and birding experience in Sri Lanka. Currently working in several projects on herpetology in the country at the HFS. Kamani Hemamala Tennekoon, Professor of Molecular Life Sciences, Institute of biochemistry, Molecular Biology and Biotechnology; University of Colombo. Current research interests include, Cancer Molecular Genetics, DNA variation, Molecular Medicine, and Medicinal Plants. Sameera R. Samarakoon (B.Sc. in Biological Science, M.Sc. in Molecular Life Sciences, Post graduate Diploma in Toxicology, PhD in Molecular Biology and Medicinal Chemistry) working as a scientific officer at the Institute of Biochemistry Molecular Biology and Biotechnology, University of Colombo, Sri Lanka. Nethu Wickramasinghe, is the Projects Co-ordinator, at the Herpetological Foundation of Sri Lanka, completed the basic degree in chemistry at the University of Delhi. A Teacher and a freelance Nature Writer, currently conducting Phylogentic Research Work, at the IBMBB, University of Colombo.
Author Contributions: LJMW was involved in designing the study, conducting the field survey, conducting museum reference work, gathering, and analyzing the data, and compiling the MS. INB was involved in field research work, writing and data compiling. DRV was involved in field research work, writing and data compiling. KHT was involved in the overall designing of the molecular study. SRS was involved in conducting and designing the molecular component of the study. NW was involved in conducting the molecular work and writing of the MS.
Acknowledgments: The authors wish to acknowledge the Biodiversity Secretariat of the Ministry of Environment, Sri Lanka for funding and collaborating the project, R.H.S.S. Samarathunga (Secretary-Ministry of Environment), Gamini Gamage (Additional Secretary-Policy Planning), Padma Abayakoon (Director), N.K.G.K. Nannawaththa (Additional Secretary-Natural Resources), R.A.R. Roopasinghe (Aditional Secretary- Administrations), Dakshini Perera, and Hasula Wickramasinghe for all the support rendered. To the Nagao Natural Environment Foundation for part funding, to the Department of Wildlife Conservation for permission granted (Permit no. WL/3/3/354), the Director General H.D. Rathnayake, U.L. Thaufeek (Deputy Director Research and Training), U.K.L. Peeris (Assitant Director Research and Training) for all the support. Chamitha for his hospitality. The authors wish to thank the Directress National Museum of Sri Lanka and staff members (Chandrika Munasinghe and Manaram de Silva), are gratefully acknowledged for their assistance in museum reference work. David Gower, Patrick Campbell, Barry Clarke, and Mark Wilkinson of the Natural History Museum, London, Alan Resetar and Kathleen Kelly of the Field Museum of Natural History are gratefully acknowledged for assisting the first author during his visit to the BMNH and FMNH, also to Roshan Rodrigo, Nalaka Aththanayaka and Naalin Perera for their hospitality extended. Sanjiv de Silva provided valuable literature, Prasanna Samarawickrama, Samantha Suranjan Fernando and Kithagama Wimalarathne thero for developing the map, relevant for this work. We wish to thank Kanishka Ukuwela for his valuable comments in the phylogenetic analysis, also Channa Bambaradeniya for his valuable comments. The authors also like to thank their colleagues at Herpetological Foundation of Sri Lanka (HFS), and the Institute of Biochemistry Molecular Biology and Biotechnology, University of Colombo for various courtesies. Asanka Abayakoon, Channaka Jayasinghe and Chamila Weerathunga for the encouragements, Sanjay Molur, B. Ravichandran and anonymous reviewers whose invaluable comments undoubtedly improved the quality of the paper. Finally to our Principal Sponsors Dilmah Conservation, for funding project activities.
Abstract: A new species of shrub frog Pseudophilautus dilmah is described from the Central Hills of Sri Lanka. This unique species is distinguished from all the other congeners from a combination of characters; snout rounded in lateral aspect, bluntly pointed in dorsal and ventral aspect, canthus rostralis rounded, vomerine teeth, lingual papilla and nuptial pads absent, dermal fringe distinct on inside of fingers III and IV, small blunt tubercles on metacarpal and ulnar folds, toes basally webbed, interorbital area smooth, upper eyelid prominent tubercles present, anterior and posterior dorsum without horny spinules but tubercles present, upper part of flank weakly granular, supratympanic fold distinct, prominent small calcar present at the distal end of the tibia, throat granular, chest and belly coarsely granular. Based on comparison of 16s rRNA gene we also show that the species is genetically distinct from other members of Pseudophilautus for which gene sequences are available. The high rate of deforestation and anthropogenic activities threaten this population in its natural habitat.
Keywords: Amphibian, biodiversity hotspot, Dilmah Shrub Frog, new taxa, Pseudophilautus dilmah.

INTRODUCTION
Sri Lanka, along with the Western Ghats of India, is one of the 34 biodiversity hotspots of the world and is credited with a rich amphibian fauna (Meegaskumbura et al. 2002; Mittermeier et al. 2004). Of recent times a large number of species has been added to the list with the total number of amphibian species adding up to 119 (see Fernando & Siriwardhane 1996; Manamendra-Arachchi & Pethiyagoda 1998, 2001, 2005; Meegaskumbura & Manamendra-Arachchi 2005; Fernando et al. 2007; Meegaskumbura et al. 2007, 2011; de Silva 2009; Wickramasinghe et al. 2012a, 2013a), while that of Pseudophilautus Laurent, 1943 now stands at 75 species (see Manamendra-Arachchi & Pethiyagoda 2005; Meegaskumbura & Manamendra-Arachchi 2005; Meegaskumbura et al. 2007, 2011, 2012; Wickramasinghe et al. 2013a). Of the 35 globally extinct amphibians prior to 2012, 21 alone were declared from Sri Lanka (Stuart et al. 2008). But after the rediscovery of three species (Wickramasinghe et al. 2012b, 2013b,c), now the total number of extinct amphibians stands at 18. Out of the predicted 18 extinct species, 17 belong to the genus Pseudophilautus (see Manamendra-Arachchi & Pethiyagoda 1998, 2005; Manamendra-Arachchi & de Silva 2004; IUCN & MENR 2007; Meegaskumbura et al. 2007; Stuart et al. 2008).
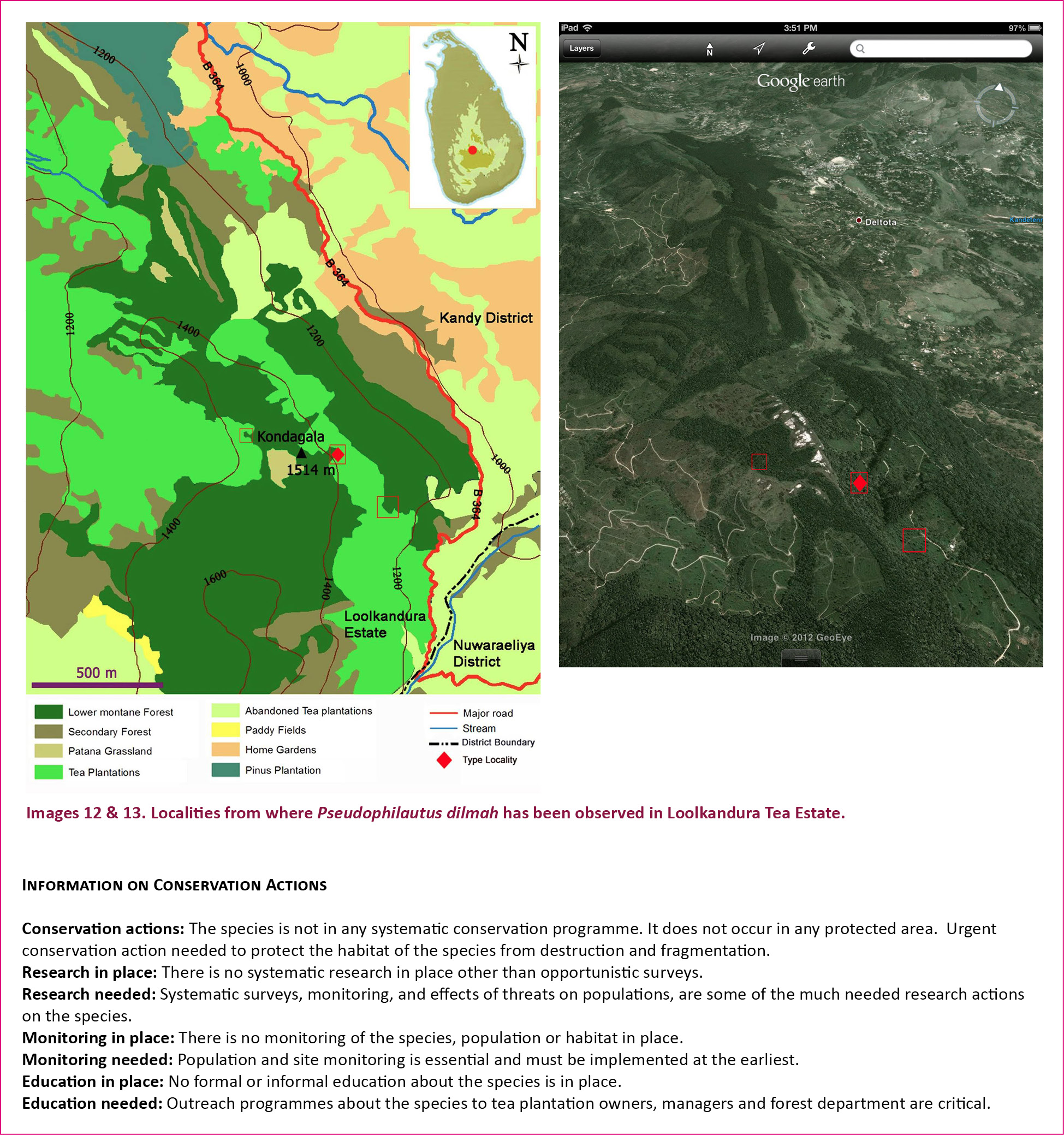
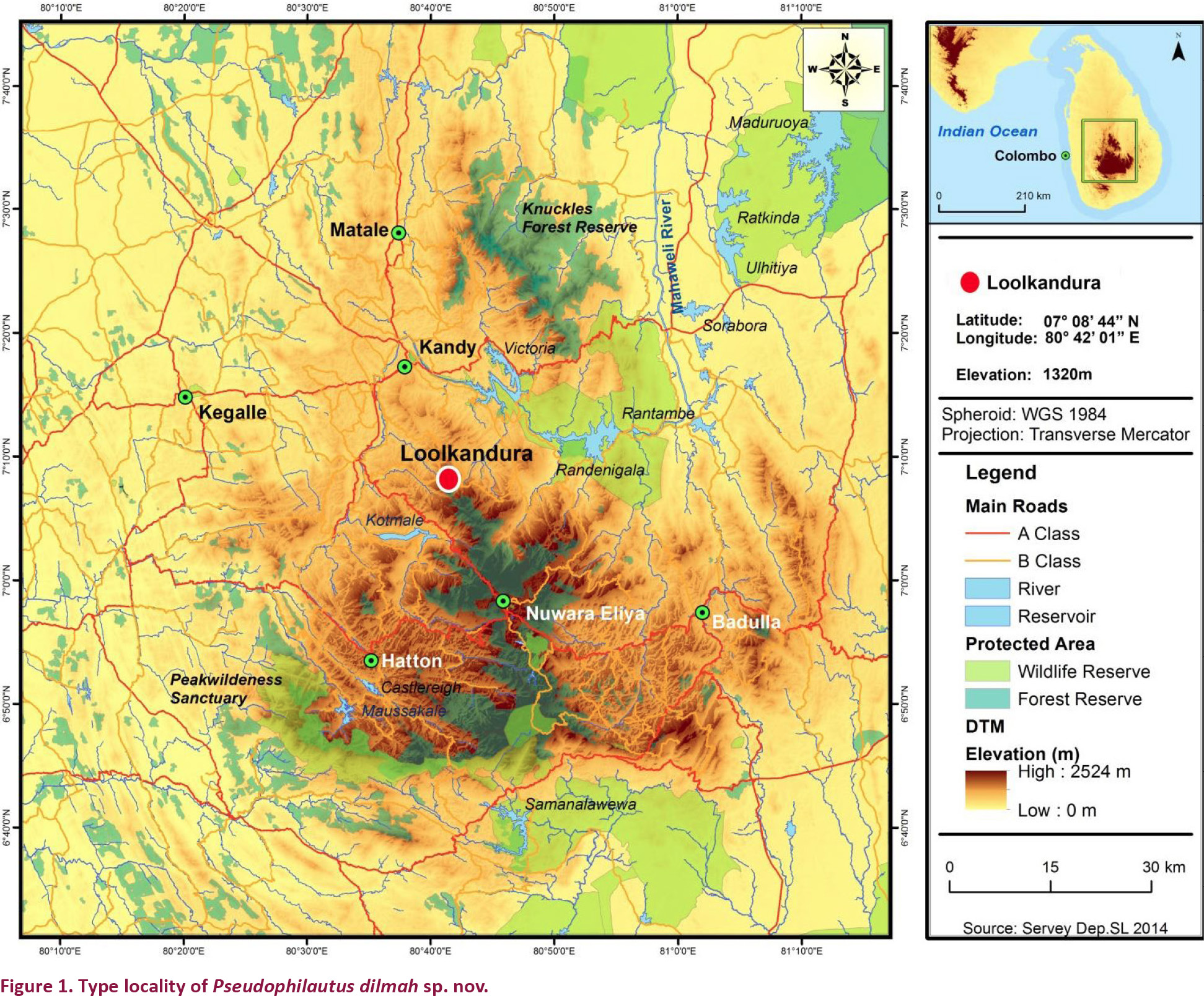
We have meticulously conducted explorations on amphibians in the last few years in the Central Hills of Sri Lanka including Peak Wilderness, Horton Plains and the Knuckles massif and subsequently describe a new species of Pseudophilautus from Loolcondera Estate (Fig. 1), adjacent to the Galaha Forest Reserve in Deltota (07017’N & 080070’E) on the border of Kandy and Nuwara-Eliya districts of Central Province.
The British colonials pronounced the Sinhala native term “Loolkandura” as “Lool-conde-ra” (Jayasuriya et al. 1993), and the English spelling has remained unchanged till date, here we shall be using the former spelling for reasons discussed later. Loolkandura and Galaha forest reserve are located in the Central Hills, which lead to Piduruthalagala, bordering the two districts of Kandy and Nuwara Eliya. Today this forest area is under the Department of Forest Conservation. The first tea plant is said to have been planted here by James Taylor in 1867, and the first tea estate established after clearing large areas of pristine forests. The tea estate is surrounded by severely fragmented and disturbed patches of pristine lower montane rain forests belonging to the Galaha Forest Reserve. The Loolkandura forest is surrounded by mostly tea estates, and a thin strip of forest connects with the Galaha Forest Reserve, while an entirely separated, large natural forest patch is located within the Loolkandura Tea Estate. The vegetation within the Loolkandura forest area can be grouped into six types, viz: natural forest, secondary forest, natural grassland, tea plantation, abandoned tea plantation, and pinus plantation.
We here describe a new species of shrub frog Pseudophilautus dilmah, from the Loolkandura forest of the Central Province of Sri Lanka. The species is reported as a result of an ongoing island-wide survey documenting the diversity of herpetofauna in Sri Lanka.
MATERIALS AND METHODS
Sampling was done mostly nocturnally, and photographs of most specimens were taken in the wild to avoid any confusion of change in colour after captivity and to avoid over collection of the same species. The specimens were photographed in life (Canon EOS 7D, Canon 100mm F/2.8 IS USM Macro Lens, Canon MT-24EX Macro Twin Lite Flash with Vello Bounce Dome Diffuser). Geographical coordinates were determined from GPS readings (Gamin eTrex Vista) at the locality. Specimens collected in the field were first fixed in 90% ethanol for two hours and stored in 70% ethanol. The material referred to is deposited in the Natural History Museum, London, UK (BMNH); the Museum of Comparative Zoology, Cambridge, USA (MCZ); the National Museum, Sri Lanka (NMSL); the Department of Wildlife Conservation, National Wildlife Research and Training Center, Girithale (DWC) (Appendix 1). The new type material discussed in this paper is also deposited in NMSL and DWC.
Morphometry
The new species was compared with all types from Sri Lanka deposited in the NMSL, and additional data were gathered from Manamendra-Arachchi and Pethiyagoda (2005), special attention was given to extinct species. The specimens formerly belonging to the Wildlife Heritage Trust (WHT) bearing WHT numbers are currently deposited in the NMSL, catalogued under the same numbers. For the description section all known species of Sri Lanka were grouped into four categories based on their snout-vent length, very small (10–15 mm), rather small (16–30 mm), moderate (31–45 mm) and large (46–60 mm). The proposed species was critically compared with species of similar morphological characters and/or with similar colouration.
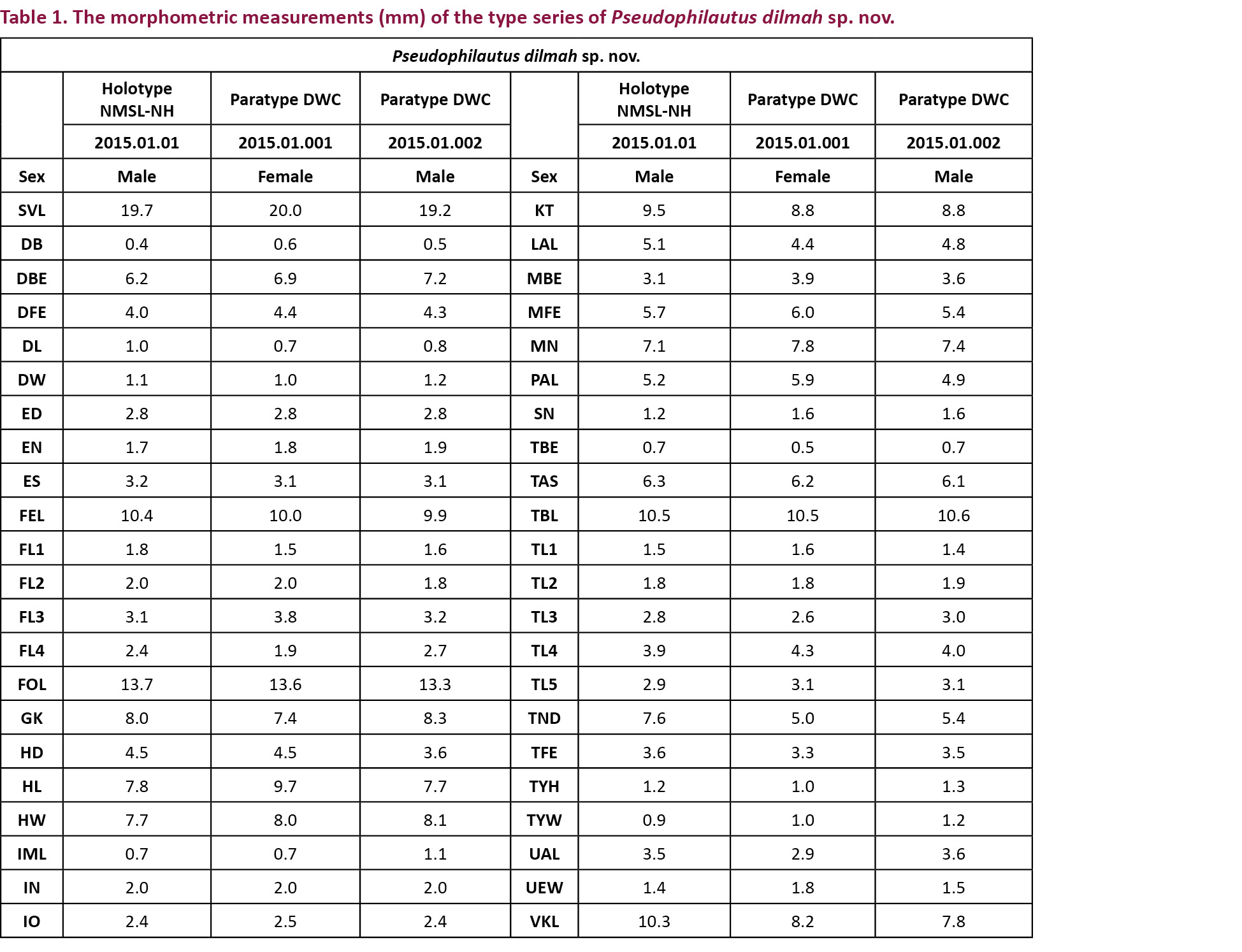
Forty-four external measurements (Table 1) of specimens were taken with a Mitutoyo digital vernier calliper to the nearest 0.1mm, and observations were made through a Leica M50 (10–40) microscope. Nomenclature of external anatomy abbreviated in the text and external measurements taken are listed alphabetically herein: maximum breadth of disk of the third finger (DB), taken from anterior to posterior edge of circum-marginal groove; distance between back of eyes (DBE), measured between posterior edge of eyes; distance between front of eyes (DFE), measured between anterior edges of eyes; length of disk of the third finger (DL), taken from anterior edge of circum-marginal groove to posterior edge of disk; width of disk of the third finger (DW), measured across the inner and outer edges of circum-marginal groove; eye diameter (ED), horizontal diameter of eye; eye to nostril distance (EN), measured between anterior most point of eye and middle of nostril; eye to snout distance (ES), measured between anterior most point of eye and tip of snout; thigh (femur) length (FEL), distance between vent and knee with both thigh and shank flexed; first finger length (FL-1), measured between proximal margin of the most proximal subarticular tubercle or crease of articulation and the tip of first finger; second finger length (FL-2), measured between proximal margin of the most proximal subarticular tubercle or crease of articulation and the tip of second finger; third finger length (FL-3), measured between proximal margin of the most proximal subarticular tubercle or crease of articulation and the tip of third finger; fourth finger length (FL-4), measured between proximal margin of the most proximal subarticular tubercle or crease of articulation and the tip of fourth finger; foot length (FOL), distance between heel and tip of fourth toe with both foot and shank flexed; groin to knee distance (GK), measured between groin and the tip of knee; head depth (HD), distance between the apex of eye and the bottom of jaw; head length (HL), distance between angle of jaws and snout tip; head width (HW), measured across angle of jaws; inner metatarsal tubercle length (IML), distance between the distal and proximal edges of the inner metatarsal tubercle; internarial distance (IN), least distance between the inner margin of nares; interorbital width (IO), least distance between the upper margins of orbits; knee-angle length (KT), distance from knee-angle to tibio-tarsal articulation; lower arm length (LAL), taken from elbow to posterior-most margin of inner palmar tubercle; mandible-back of eye distance (MBE), distance between angle of jaws and posterior-most point of eye; mandible-front of eye distance (MFE), taken as distance between angle of jaws and anterior-most point of eye; mandible-nostril distance (MN), taken as distance between angle of jaws and middle of nostril; palm length (PAL), taken from posterior-most margin of inner palmar tubercle to tip of disk of third finger; snout-nostril distance (SN), taken as distances between middle of nostril and tip of snout; snout-vent length (SVL), measured from tip of snout to vent; tympanum-back of eye distance (TBE), distance between anterior-most point of inner margin of inner rim of tympanum and posterior most point of eye; tarsal length (TAS), measured between the tibio-tarsal articulation and anterior edge of inner metatarsal tubercle; tibia length (TBL), distance between knee and heel with both shank and foot flexed; first toe length (TL-1), measured between proximal margin of the most proximal subarticular tubercle or crease of articulation and the tip of first toe; second toe length (TL-2), measured between proximal margin of the most proximal subarticular tubercle or crease of articulation and the tip of second toe; third toe length (TL-3), measured between proximal margin of the most proximal subarticular tubercle or crease of articulation and the tip of third toe; fourth toe length (TL-4), measured between proximal margin of the most proximal subarticular tubercle or crease of articulation and the tip of fourth toe; fifth toe length (TL-5), measured between proximal margin of the most proximal subarticular tubercle or crease of articulation and the tip of fifth toe; tympanum-nostril distance (TND), distance between anterior-most point of inner margin of inner rim of tympanum and middle point of nostril; tympanum-front of eye distance (TFE), taken as distance between anterior-most point of inner margin of inner rim of tympanum and anterior-most point of eye; tympanum height (TYH), vertical diameter of the inner rim; tympanum width (TYW), horizontal diameter of the inner rim; upper arm length (UAL), distance between axilla and elbow; upper eyelid width (UEW), measured from bony edge of supraorbital to outer edge of upper eyelid; vent to knee-angle length (VKL), distance measured from knee-angle to vent.
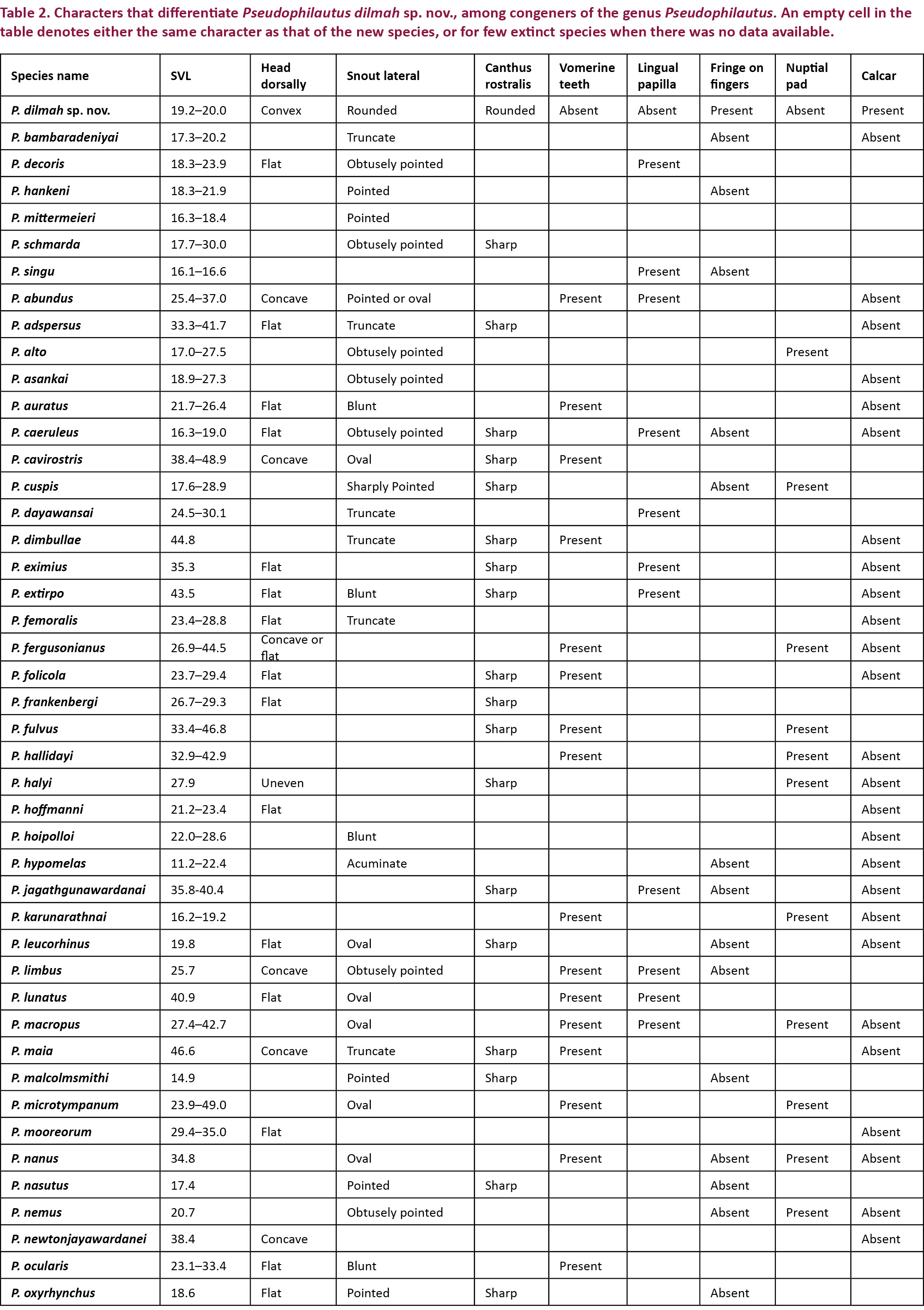
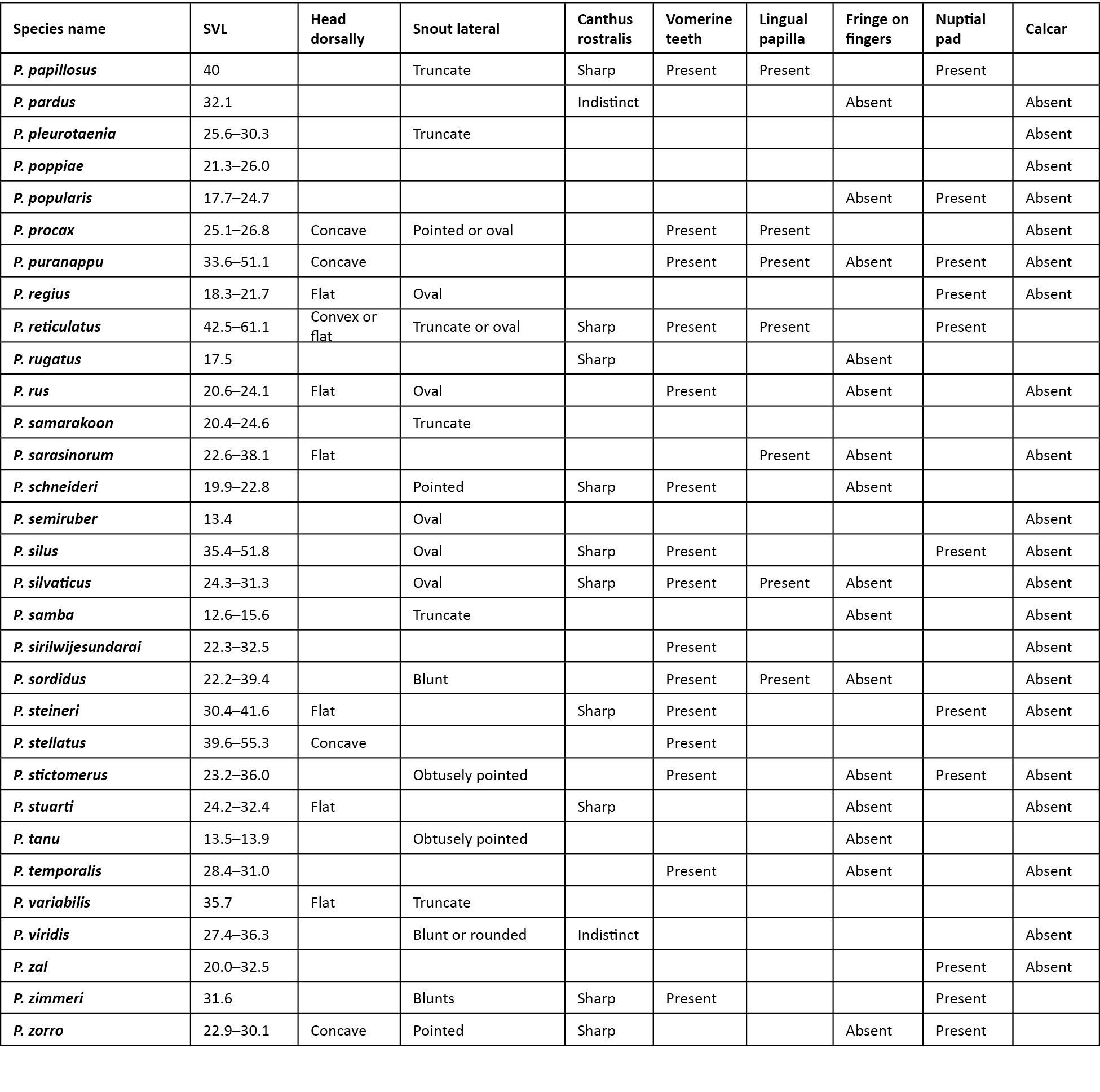
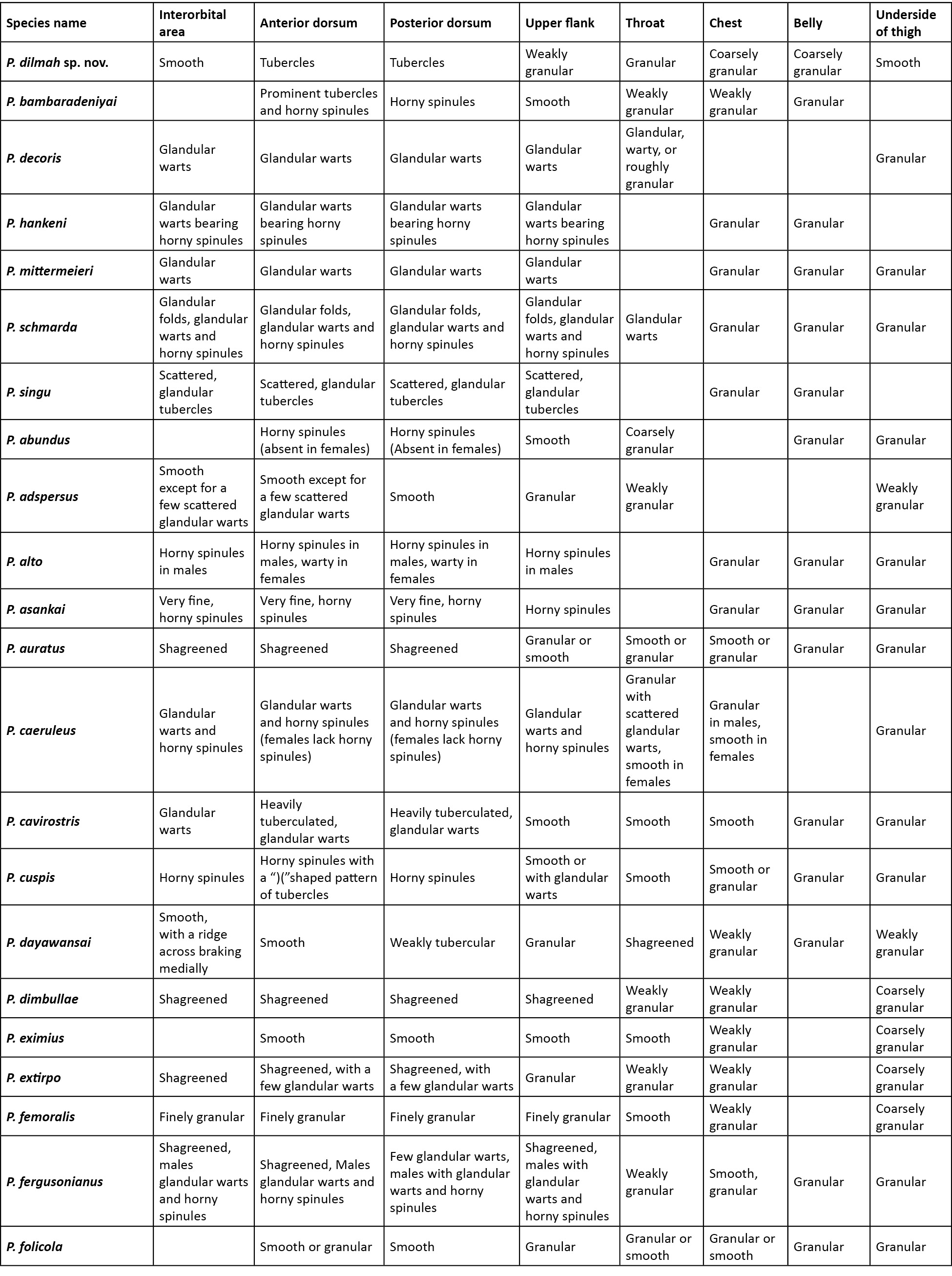
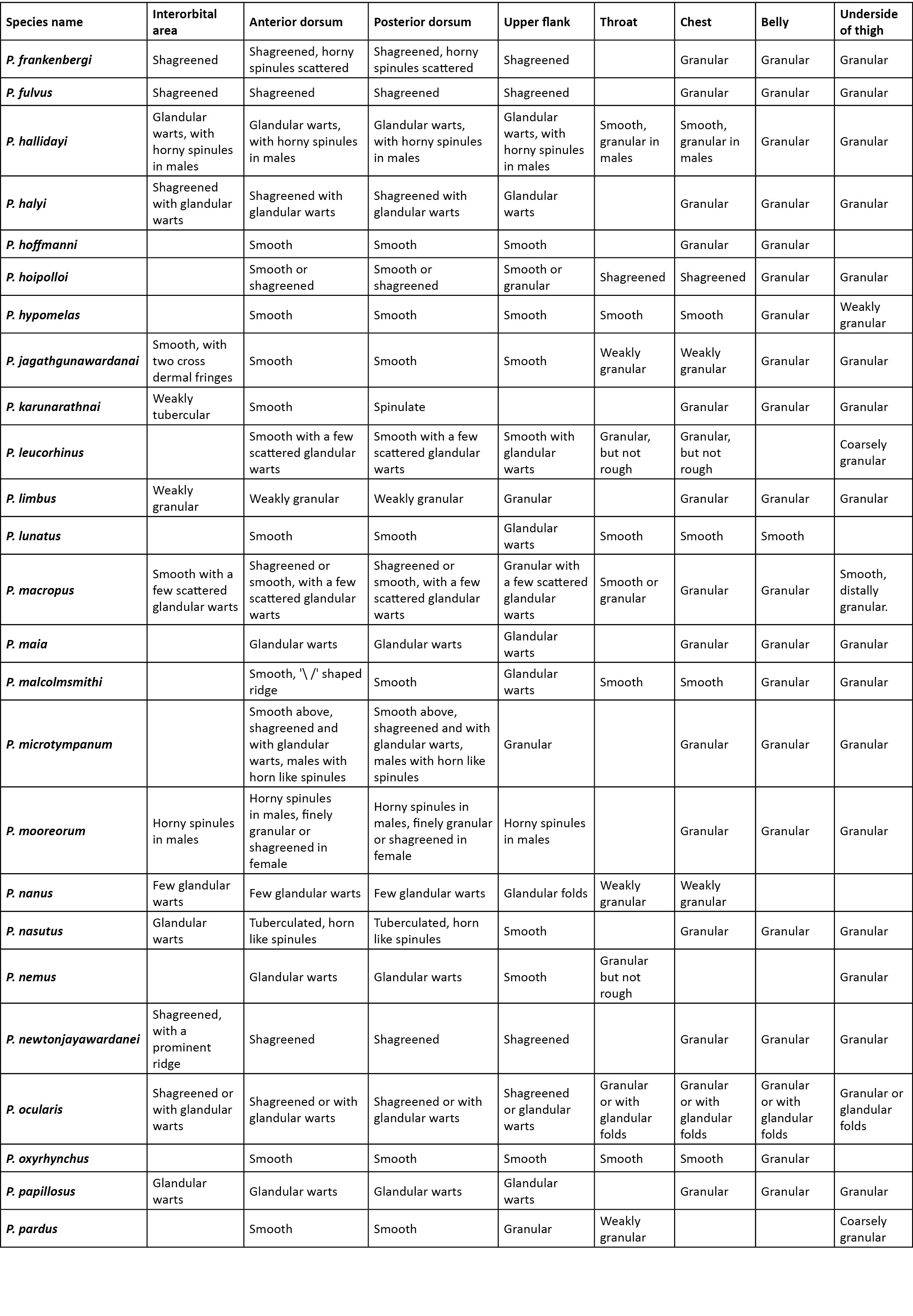
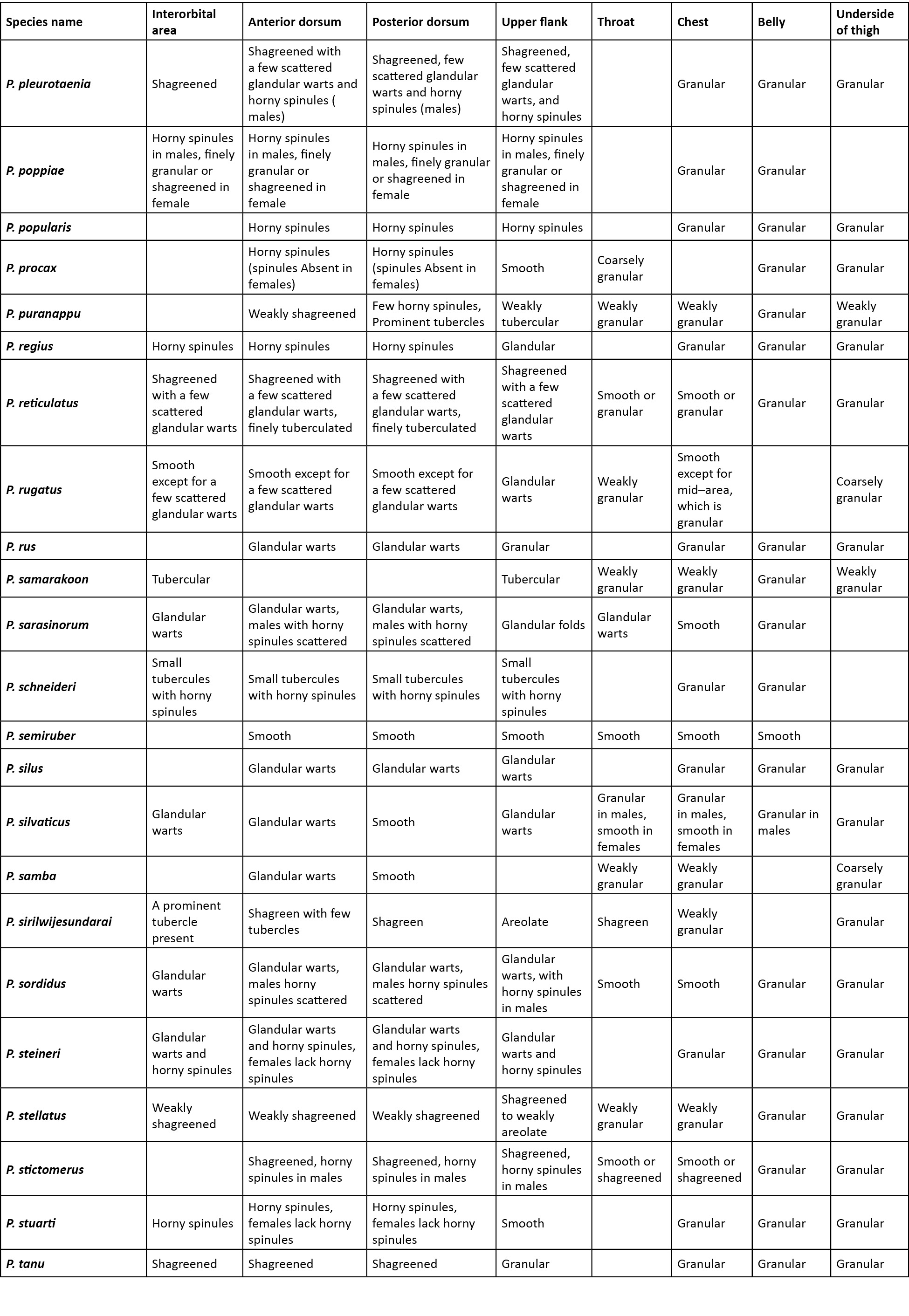
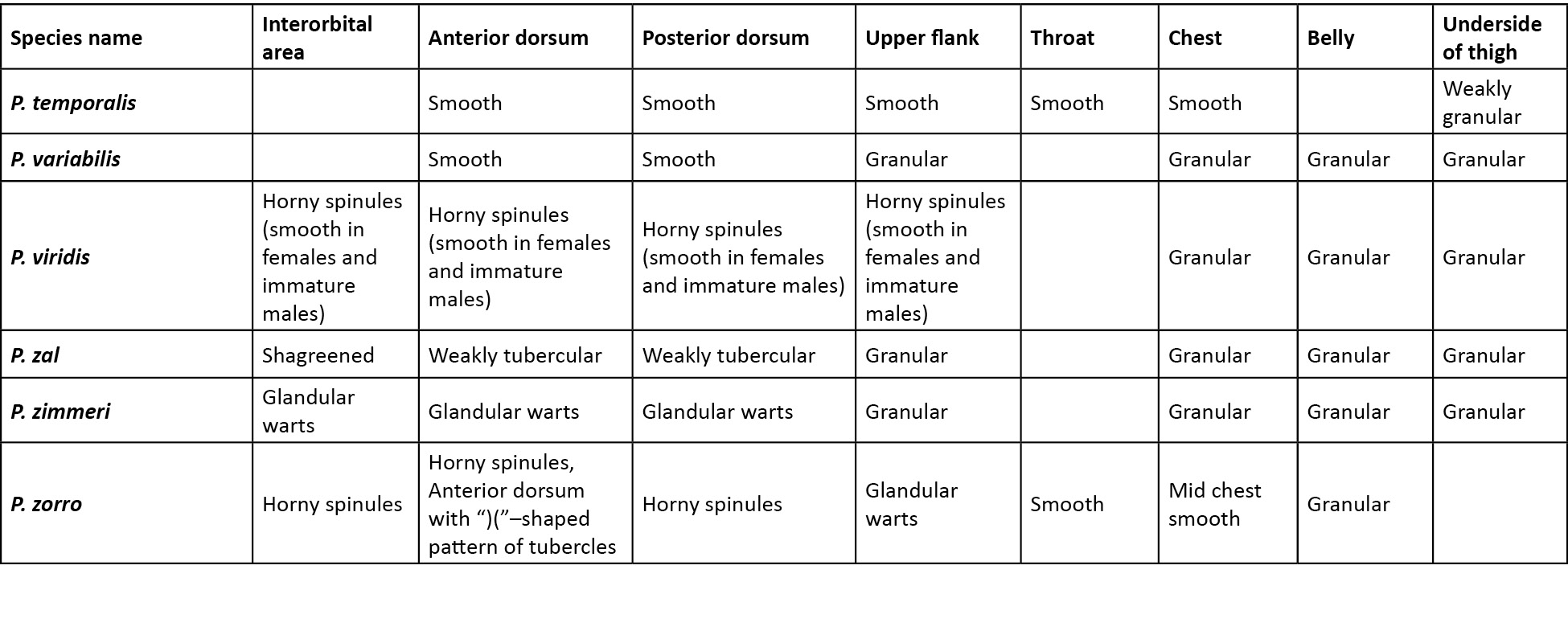
Comparison was carried out amongst congeners which closely resembled the new species. In the comparison tables for morphological characters (Table 2), and characters in the skin (Table 3), an empty cell in the table denotes either the same character as that of the new species, or for few extinct species when there was no data available.
Additional data for comparison were collected from Meegaskumbura & Manamendra-Arachchi (2005, 2011); Meegaskumbura et al. (2009); Pethiyagoda & Manamendraarachchi (2005); Wickramasinghe et al. (2013a).
Molecular phylogentic analysis
The phylogenetic relationships of the newly described species to other species of the genus Pseudophilautus in Sri Lanka, for which data is available, were estimated using the mitochondrial 16S ribosomal RNA region. Whole cellular DNA was extracted from ethanol preserved muscle tissue sample from the specimen number DWC 2015.01.001 of the new species and NMSL 2013.01.01 NH of P. bambaradeniyai. The tissue sample was digested at 550C using STE buffer (0.1 M NaCl, 0.01 M TRIS, EDTA 0.001 M and 25 μl of 10% SDS) with 10μl of 20 mg/ml Proteinase K. Final extraction was carried out using the phenol-chloroform method. Polymerase chain reaction (PCR) amplifications of the 16S ribosomal RNA region were carried out in 25 μl reactions using the forward primer- 5’-GCCTGTTTATCAAAAACAT-3’ and reverse primer 5’-CCGGTCTGAACTCAGATCACGT-3’ (Frost et al. 2006) with Promega GoTaq DNA polymerase enzyme in an Eppendorf Master cycler. The thermal profile was an initial denaturation step for seven minutes at 940C, and 35 cycles of denaturation for 40 seconds at 940C, annealing for 30 seconds at 520C and extension for 30 seconds at 720C, followed by final extension for 10 min at 720C. Amplified DNA fragments were purified using the ‘Promega Wizard Gel and PCR clean up’ system. Purified PCR products were sequenced using a BigDye Terminator sequencer v. 3.0 (Applied BioSystems, CA, USA) in 8 μl reactions. The sequence chromatograms were visualized in a Genetic Analyzer 3500 Dx (Applied Biosystems, CA, USA).
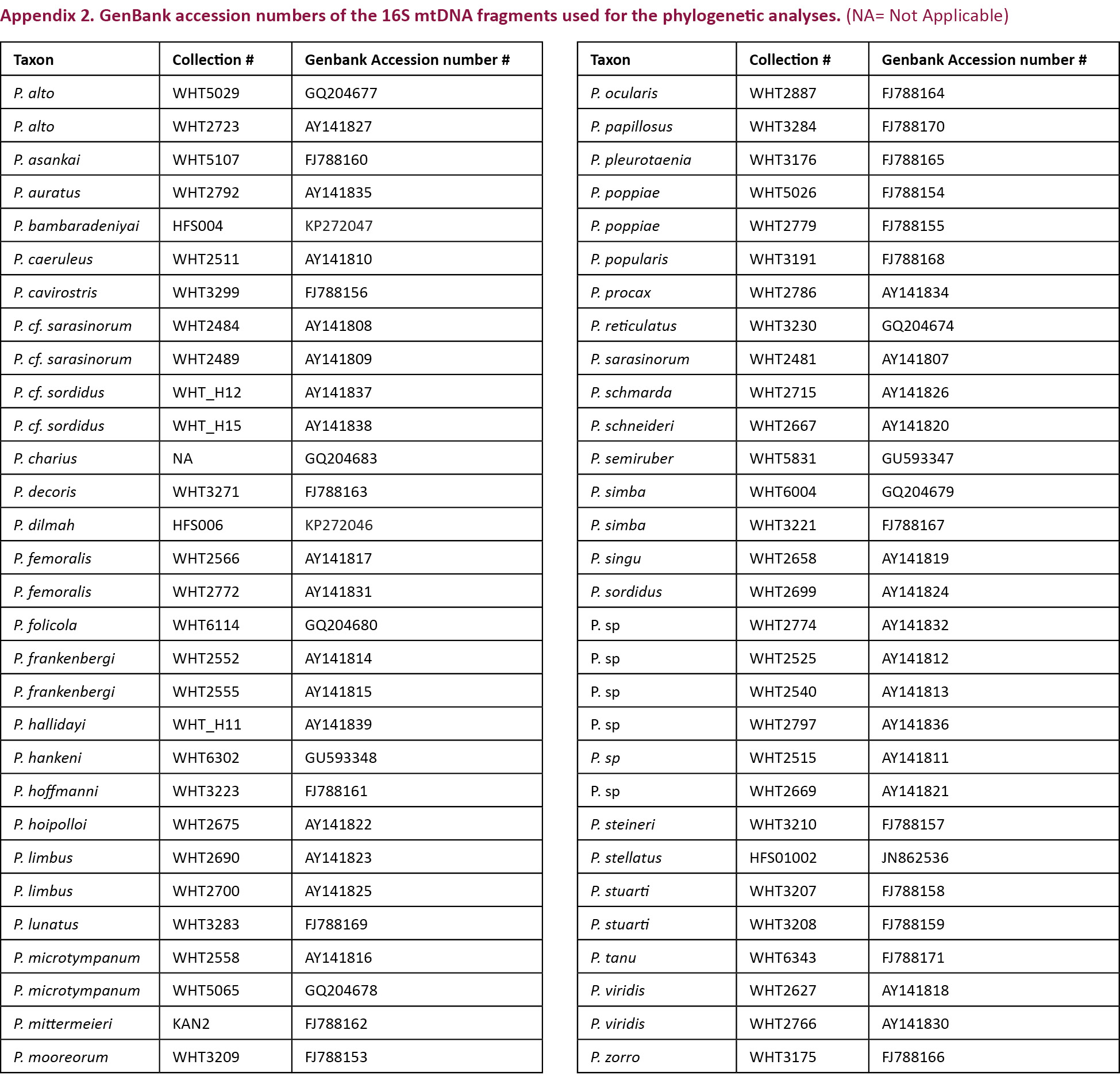
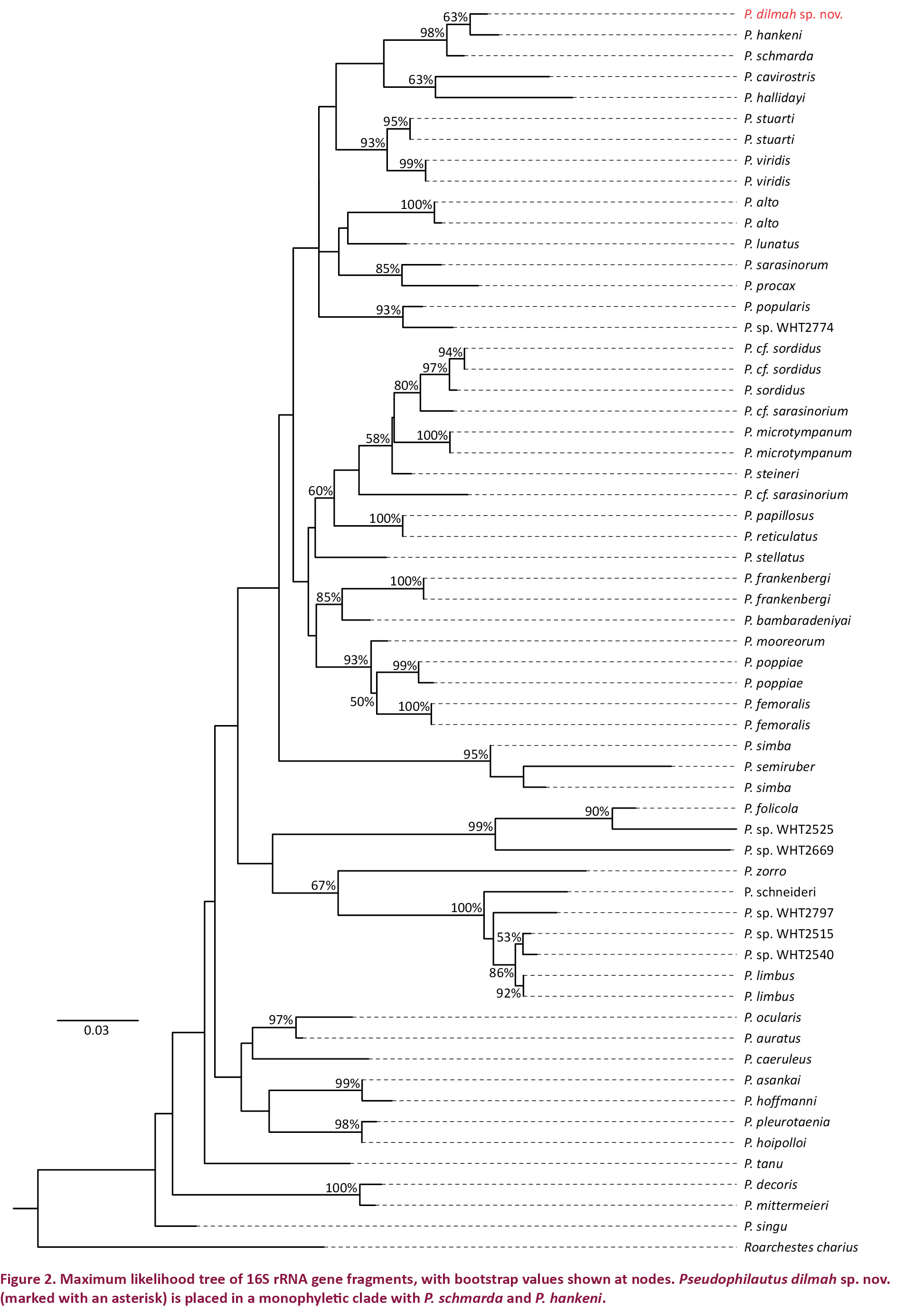
The newly generated sequences have been deposited in GenBank under the accession numbers KP272046 and KP272047. Additional sequences of the Sri Lankan Pseudophilautus species were obtained from GenBank (Appendix 2). Gene sequences were aligned using MUSCLE (Edgar 2004). Molecular phylogenetic analysis was performed using MEGA 6 (Tamura et al. 2013). The best fit model for nucleotide substitution was selected from 24 models using MEGA 6 based on the minimum Bayesian Information Criterion (BIC) value (Schwarz 1978; Nei & Kumar 2000). Best fit model was used for computing the pairwise distances between the sequences and for maximum likelihood analysis. Maximum likelihood tree was rooted with Raorchestes charius, an Indian species that represent Raorchestes, the sister group to the Sri Lankan genus Pseudophilautus. Tree was drawn using FigTree v1.4.2 (Morariu et al. 2009).
RESULTS
Pseudophilautus dilmah sp. nov.
(Figs. 1–2; Images 1–4; Table 1)
urn:lsid:zoobank.org:act:C7C025BE-35ED-4519-A8BD-B36BF82B8B6C
Holotype: NMSL 2015.01.01 NH, 01.v.2011, adult male 19.7mm SVL (Image 1), Loolkandura (Lulcondera), Kandy District, Central Province, Sri Lanka (07008’44.96”N & 080042’01.13”E; 1324m) coll. L.J.M.W. & I.N.B.
Paratypes: DWC 2015.01.001, adult female, 20.0mm SVL (Image 2); DWC 2015.01.002, adult male, 19.2mm; the same date, locality and collectors.
Diagnosis: Body rather small size (SVL 19–20 mm). Snout rounded in lateral aspect, bluntly pointed in dorsal and ventral aspect. Canthus rostralis rounded. Vomerine teeth, lingual papilla and nuptial pads absent. Dermal fringe distinct on inside of fingers III and IV, small blunt tubercles on metacarpal and ulnar folds. Toes basally webbed. Interorbital area smooth. Upper eyelid prominent tubercles present. Anterior and posterior dorsum without horny spinules but tubercles present. Upper part of flank weakly granular. Supratympanic fold distinct. Prominent small calcar present at the distal end of the tibia. Throat granular, chest and belly coarsely granular.

Description of holotype
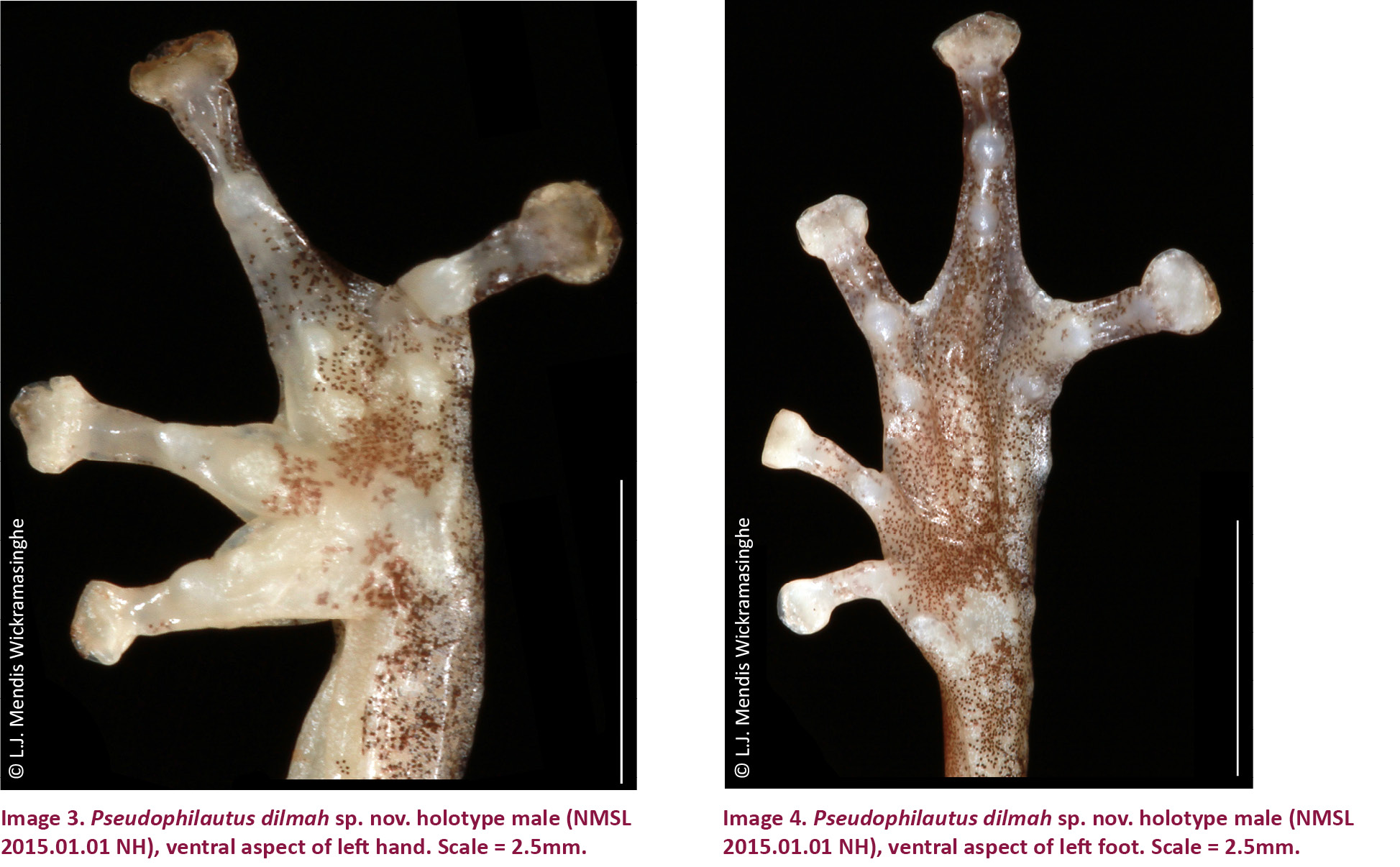
Body small size (SVL 19.7mm), elongate (SVL/HW 2.6). Head small (HL/SVL 0.4), about as wide as long (HW/HL 1.0), convex above. Snout rounded in lateral aspect, bluntly pointed in dorsal and ventral aspect (ES/DFE 0.8, SN/IN 0.6), its length longer than horizontal diameter of eye (ES/ED 1.2). Internasal space and loreal region concave. Canthus rostralis rounded. Interorbital space convex, larger than upper eyelid (IO/UEW 1.7), and internasal distance (IN/UEW 1.4); distance between front of eyes 3/5th the distance between back of eyes (DBE/DFE 1.6). Nostrils oval, without flap of skin laterally, closer to tip of snout than to eye (SN/EN 0.7). Pupil horizontally elliptical. Tympanum distinct, oval (TYH/TYW 1.3), smaller than half eye diameter (TYH/ED 0.4), tympanum-eye distance 3/4th the tympanum width (TBE/TYW 0.8). Pineal ocellus, vomerine teeth, and lingual papilla absent. Arm short, thin (LAL/FEL 0.5, UAL/FEL 0.3). Lower arm as long as palm length (LAL/PAL 1.0), longer than upper arm (LAL/UAL 1.4). Fingers thin, 3rd finger the longest (Image 3). Relative length of fingers I < II < IV < III (FL-1/FL-3 0.6, FL-2/FL-3 0.6, FL-4/FL-3 0.8), (Table 1). Tips of fingers rounded enlarged, discs present on all fingers, with distinct basal and circum marginal grooves. Dermal fringe distinct on inside of fingers III and IV, small blunt tubercles on metacarpal and ulnar folds. Rudimentary webbing present. Distal subarticular tubercles present in all fingers; prominent, rounded, single. Penultimate subarticular tubercles present in third and fourth fingers, rounded, single. Inner palmar tubercle indistinct, single, oval, larger than outer palmer tubercle. Nuptial pads absent. Outer palmar tubercle indistinct, single, oval, smaller than distal subarticular tubercles. Few supernumerary tubercles present on palm. Prepollex absent. Femur longer than fourth toe length (FEL/TL-4 2.7), foot length longer than thigh (FOL/FEL 1.3), tarsus shorter than tibia length (TAS/TBL 0.6). Toes thin, 4th toe being the longest (Image 4). Relative length of toes I < II < III < V<IV (TL-1/TL-4 0.4, TL-2/TL-4 0.5, TL-3/TL-4 0.7, TL-5/TL-4 0.7). Tips of toes rounded, enlarged, discs present on all toes with distinct basal and circum marginal grooves. All toes basally webbed. Dermal fringe present on all toes, fringe along postaxial edge of toe V, and metatarsal fold, small undulating tubercles present on outer edge of tarsal fold. Distal subarticular tubercles prominent, rounded and single, present on all toes, larger than other tubercles (except metatarsal tubercles). Penultimate subarticular tubercles present in III, IV and V toes, rounded, and single. Anti penultimate subarticular tubercles fourth and fifth toes, indistinct, rounded, and single. Few supernumerary tubercle present, indistinct. Inner metatarsal tubercle oval prominent and large, its length half in length of toe I (IML/TL-1 0.5). Outer metatarsal tubercle present, small and indistinct.
Skin of dorsal and lateral body
Dorsal and lateral regions of snout smooth. Prominent tubercles between eyes and on upper eyelid. Side of head smooth. Median dermal ridge absent. Anterior and posterior part of back with tubercles, without horny spinules. Upper part of flank weakly granular. Lower part of flank granular. Latero-dorsal folds, lateral line system, fejervarya line, cephalic ridges, and co-ossified skin absent. Supratympanic fold distinct. Upper arm, lower arm, and foot smooth. Anterior, dorsal, and posterior part of thigh smooth. Leg, and tarsus smooth with few tubercles. Prominent small calcar present at the distal end of the tibia. Hand smooth.
Skin of ventral part
Throat granular, margin of throat smooth with prominent tubercles. Chest and belly coarsely granular. Upper arm granular. Lower arm smooth. Thigh smooth. Leg smooth. Tarsus smooth. Vent anteriorly smooth, either side weakly granular. Macroglands absent.
Colour in life
Dorsum cream with light brown patches, dark brown cross band between eyes, pair of dark brown dots placed behind the cross band, a pair of broad light brown longitudinal bands ending at the sacrum, dark brown blotches on groin, lateral body lighter with a light olive greenish tinge; limbs cream, fore limbs, hind limbs, fingers and toes with light brown cross bands; ventral side belly off white with light brown blotches, chest off white with few blotching, throat uniform off white, hands, feet and webbing lighter (Image 1).
Colour in alcohol
Colour pattern remains with a little darkening, ventral side off white with dark brown blotching.
Colour variation
Dorsal colour varies ranging to a lighter brownish tinge, and at the ventral side belly shows prominent dark blotches.
Etymology
The species epithet dilmah is named after Dilmah Conservation, for its dedicated efforts to biodiversity conservation on the Island. Dilmah is treated as an invariable noun in apposition to the generic name.
Suggested vernacular names
Dilmah panduru madiya, and Dilmah Shrub Frog in Sinhala, and English respectively.
Molecular analysis
The final alignment consisted of a total of 590 base pairs (bp). Model test identified Tamura & Nei (1993) nucleotide substitution model with gamma distribution (BIC = 7964.99, lnL = -3360.55, G = 0.26) as the best-fit model. Maximum likelihood analysis of the 16S rRNA fragment (Fig. 2) places Pseudophilautus dilmah sp. nov. in a monophyletic group along with P. schmarda (Kelaart 1854) and P. hankeni Meegaskumbura & Manamendra-Arachchi 2011. Pairwise genetic distance between P. dilmah sp. nov. and P. schmarda was 1.9% while it was 1.6% between the former and P. hankeni. There was a 10% divergence in the 16s rRNA gene between P. dilmah sp. nov. and P. bambaradeniyai Wickramasinghe et al. 2012, the morphologically most closely related species.
Comparison
A critical comparison was made amongst all congeners, and Table 2 provides discriminating morphological characters against all congeners, while Table 3 provides discriminating characters of the skin and it was evident that Pseudophilautus dilmah sp. nov., resembles P. bambaradeniyai (Image 5), due to similar colour pattern, comparable adult body size, and because both species are found in the Central Hills. Pseudophilautus dilmah, can be easily differentiated from P. bambaradeniyai by the absence of horny spinules on the anterior dorsum (vs present), snout rounded in lateral aspect (vs truncate), fringe on fingers present (vs absent), and calcar present (vs absent).
Although P. dilmah sp. nov., is genetically most closest to P. hankeni (Image 6) and P. schmarda (Image 7), with genetic distances of 1.6% and 1.9% respectively, the species is morphologically distinct and can be separated by the presence of the following combination of characters: from P. hankeni, by snout in lateral aspect rounded (vs pointed), fringe on fingers present (vs absent), interorbital area smooth (vs glandular warts bearing horny spinules), dorsum having tubercles (vs glandular warts bearing horny spinules), upper flank weakly granular (vs glandular warts bearing horny spinules), chest and belly coarsely granular (vs granular); from P. schmarda by snout rounded in lateral aspect (vs obtusely pointed), canthus rostralis rounded (vs sharp), interorbital area smooth (vs glandular folds, glandular warts and horny spinules), dorsum having tubercles (vs glandular folds, glandular warts and horny spinules), upper flank weakly granular (vs glandular warts bearing horny spinules), throat granular (vs glandular warts), chest and belly coarsely granular (vs granular), underside of thigh smooth (vs granular).
To a lesser extent P. dilmah may be confused with P. decoris (Manamendra-Arachchi & Pethiyagoda 2005), P. mittermeieri (Meegaskumbura & Manamendra-Arachchi 2005), and P. singu (Meegaskumbura et al., 2009), because of similar morphological characters, hence these species were included at the beginning of the comparison tables.

DISCUSSION
Pseudophilautus dilmah sp. nov., appears to be more resembling to P. bambaradeniyai, (Fig. 2) than to any other species of the same genus. But considering molecular evidences P. bambaradeniyai has been placed in a well separated clade with P. frankensbergi (Meegaskumbura & Manamendra-Arachchi 2005) with a pairwise genetic distance of 10%, which suggests a species level divergence. Although they were both found from the Central Hills they are allopatric. Loolkandura the type locality of P. dilmah sp. nov., is positioned towards the northern tip of the Central Hills where as Peak Wilderness the type locality of P. bambaradeniyai, and the only locality it is found in, is positioned towards south-west of Loolkandura (Fig. 1).
Although P. dilmah sp. nov., is genetically most closest to P. hankeni and P. schmarda, with genetic distances of 1.6% and 1.9%, respectively, the species is morphologically distinct and can be separated by the characters mentioned in the comparison. Furthermore P. hankeni is distributed in the Knuckles massif which is geographically well separated, and although P. schmarda is distributed in the Central Hills they are allopatric (Fig. 1). Although 3% genetic distance is a good indication Vences et al. (2005) mention that interspecies genetic distances could be from 1% to 10% and allopatric species are known with less than 3% differences. The pairwise distance for P. pleurotaenia (Boulenger 1904) and P. hoipolloi (Manamendra-Arachchi & Pethiyagoda 2005), for P. asankai (Manamendra-Arachchi & Pethiyagoda 2005) and P. hoffmanni (Meegaskumbura & Manamendra-Arachchi 2005), for P. decoris and P. mittermeieri, were 0.5%, 1%, and 1.4%, respectively. All of which are considered valid morphologically yet have a lower genetic distance than 1.6% for P. dilmah and P. hankeni, and 1.9% for P. dilmah and P. schmarda, hence our current genetic distances from its sister taxa suggests species level divergence.
Although to a lesser extent considering morphological characteristics P. dilmah may be confused with P. decoris; distributed in the Rakwana range, pairwise genetic distance of 18.3%; with P. mittermeieri; distributed in the lowland rain forests to lower montane rain forests, pairwise genetic distance of 17.3%, and with P. singu; distributed in the lowland rain forests, pairwise genetic distance of 10.1%, considering their geographic distribution and molecular divergence the new species is well separated from all the above three species.
The natural forest cover in the type locality is fast declining mainly due to encroachment by tea plantations (Images 8–10), which has evidently led to forest fragmentation (Image 11). In order to protect the species from habitat loss, the few remaining areas which is home to the species needs legal protection. It is hoped with the discovery of the species in this locality the authorities will take necessary measures to protect the natural forest cover which is home to the newly described species P. dilmah.


Proposed conservation status
The species was recorded from an elevation of about 1300m, and commonly seen perched on 1–2m high bushes. The specimens were found in natural forest cover as well as in the disturbed areas, with no canopy cover and in areas with regenerated forest covers. The area which the species was reported is currently under severe anthropogenic pressures. Since the colonial period the area was subjected to clearing of pristine forest covers for coffee, cinchona and later tea and cardamom plantations. A few remnant patches of unique “mid elevation lower montane forest”, which shows the mixed characteristics of montane forests and lower montane forests, exist in some areas. Especially, natural forests remain only in the tough terrains where colonials could not spread the plantations and most of the stream banks are also severely affected due to encroaching tea cultivations (Images 8–11).
The proposed conservation status for the species according to the IUCN Red List Criteria can be considered Critically Endangered (criteria B1ab(iii)). Since the extent of occurrence is <100km2, is recorded from a single location, and their habitats under sever threat.
REFERENCES
de Silva, A. (2009). Amphibians of Sri Lanka; A Photographic Guide to Common Frogs, Toads and Caecilians. Published by author, 250pp.
Dinerstein, E. & E.D. Wickramanayake (1993). Beyond “Hotspots”: How to prioritize investments to conserve biodiversity in the Indo-Pacific region. Conservation Biology 3(1): 53–65.
Edgar, R.C. (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research 32: 1792–1797; http://dx.doi.org/10.1093/nar/gkh340
Fernando S.S., L.J.M. Wickramasinghe & R.K. Rodirigo (2007). A new species of endemic frog belonging to genus Nannophrys Gunther, 1869 (Anura: Dicroglossinae) from Sri Lanka. Zootaxa 1403: 55–68.
Fernando, P. & M. Siriwardhane (1996). Microhyla karunaratnei (Anura: Microhylidae), a new species of frog endemic to Sri Lanka. Journal of South Asian Natural History 2: 135–142.
Frost, D.R., T. Grant, J.N. Faivovich, R.H. Bain, A. Haas, C.L.F.B. Haddad, R.O. De Sa, A. Channing, M. Wilkinson, S.C. Donnellan, C.J. Raxworthy, J.A. Campbell, B.L. Blotto, P. Moler, R.C. Drewes, R.A. Nussbaum, J.D. Lynch, D.M. Green & W.C. Wheeler (2006). The amphibian tree of life. Bulletin of the American Museum of Natural History 297: 1–370; http://dx.doi.org/10.1206/0003-0090(2006)297[0001:TATOL]2.0.CO;2
Gunawardene, N.R., A.E.D. Daniels, I.A.U.N. Gunatilleke, C.V.S. Gunatilleke, P.V. Karunakaran, G.K. Nayak, S. Prasad, P. Puyravaud, B.R. Ramesh, K.A. Subramanian & G. Vasanthy (2007). A brief overview of the Western Ghats - Sri Lanka biodiversity hotspot. Current Science 93(11): 1567–1572.
IUCN Sri Lanka & Ministry of Environment and Natural Resources Sri Lanka (2007). The 2007 National Red List of Threatened Fauna and Flora of Sri Lanka. IUCN Sri Lanka and Ministry of Environment and Natural Resources Sri Lanka, 148pp.
Jayasuriya, A.H.M., A.M. Greller, S. Balasubramaniam, C.V.S. Gunatilleke, I.A.U.N. Gunatilleke & M.D. Dassanayake (1993). Phytsociological studies of mid-elevational (lower montane) evergreen mixed forests in Sri Lanka. Proceeding of International and Interdisciplinary Symposium. Ecology and Landscape Management in Sri Lanka 79–94pp.
Manamendra-Arachchi, K. & A. de Silva (2004). Adenomus kandianus. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.2. <www.iucnredlist.org>. Downloaded on 18 January 2013.
Manamendra-Arachchi, K. & R. Pethiyagoda (1998). A synopsis of the Sri Lankan Bufonidae (Amphibia: Anura) with description of new species. Journal of South Asian Natural History 3: 213–248.
Manamendra-Arachchi, K. & R. Pethiyagoda (2001). Polypedates fastigo, a new tree frog (Ranidae: Rhacophorinae) from Sri Lanka. Journal of South Asian Natural History 5(2): 191–199.
Manamendra-Arachchi, K. & R. Pethiyagoda (2005). The Sri Lankan shrub frogs of the genus Philautus Gistel, 1848 (Ranidae: Rhacophorinae), with description of 27 new species. The Raffles Bulletin of Zoology 12: 163–303.
Meegaskumbura, M. & K. Manamendra-Arachchi (2005). Description of eight new species of shrub-frogs (Ranidae: Rhacophorinae: Philautus) from Sri Lanka. The Raffles Bulletin of Zoology 12: 305–338.
Meegaskumbura, M., F. Bossuyt, R. Pethiyagoda, K. Manamendra-Ararchchi, M. Bahir, M.C. Milinkovitch & C.J. Schneider (2002). Sri Lanka: an amphibian hotspot. Science 298: 379.
Meegaskumbura, M., K. Manamendra-Arachchi, C.J. Schneider & R. Pethiyagoda (2007). New species amongst Sri Lanka’s extinct shrub frogs. Zootaxa 1397: 1–15.
Meegaskumbura, M., K. Manamendra-Arachchi, G. Bowatte & S. Meegaskumbura (2012). Rediscovery of Pseudophilautus semiruber, a diminutive shrub frog (Rhacophoridae: Pseudophilautus) from Sri Lanka. Zootaxa 3229: 58–68.
Meegaskumbura, M., S. Meegaskumbura, G. Bowatte, K. Manamendra-Arachchi, R. Pethiyagoda, J. Hanken & C.J. Schneider (2011). Taruga (Anura: Rhacophoridae), a new genus of foam-nesting tree frogs endemic to Sri Lanka. Ceylon Journal of Science (Biological Sciences) 39(2): 75–94.
Mittermeier, R.A., P.R. Gil, M. Hoffmann, J. Pilgrim, T. Brooks, C.G. Mittermeier, J. Lamoreux & G.A.B.D. Fonseca (2004). Hotspots Revisited Earth’s Biologically Richest and Most Endangered Terrestrial Eco-regions. Conservation International. Sierra Madre and University of Virginia, 390pp.
Moorman, F.R. & C.R. Panabokke (1961). Soils of Ceylon. Tropical Agriculturist 117: 4–65.
Morariu, V. I., B.V. Srinivasan,V.C. Raykar, R. Duraiswami & L.S. Davis (2009). Automatic online tuning for fast Gaussian summation, pp. 1113–1120. In: Advances in Neural Information Processing Systems. http://www.umiacs.umd.edu/~morariu/publications/MorariuFigtreeNIPS08_preprint.pdf. Accessed 10 February 2015.
Nei, M. & S. Kumar (2000). Molecular Evolution and Phylogenetics. Oxford University Press, New York, 333pp.
Schwarz, G. (1978). Estimating the dimension of a model. The Annals of Statistics 6(2): 461–464.
Sri Lanka Tea Board (2003). Sri Lanka Tea Board, POB 1750, Colombo, Sri Lanka. http://www.pureceylontea.com/history.htm. Accessed 10 March 2012.
Stuart, S.N., M. Hoffmann, J.S. Chanson, N.A. Cox, R.J. Berridge, P. Ramani & B.E. Young (2008). Threatened Amphibians of the World. Lynx Editions, 776pp.
Tamura, K., G. Stencher, D. Peterson, A. Filipski & S. Kumar (2013). MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Molecular Biology and Evolution 30(12): 2725–2729; http://dx.doi.org/10.1093/molbev/mst197
Vences, M., M. Thomas, A. van der Meijden, Y. Chiari & D.R. Vieites (2005). Comparative performance of the 16S rRNA gene in DNA barcoding of amphibians. Frontiers in Zoology 2(1): 1–12; http://dx.doi.org/10.1186/1742-9994-2-5
Whitmore, T.C. (1984). Tropical Rain Forests of The Far East (2nd Edition). Oxford University Press, Oxford, xvi+352pp.
Wickramasinghe, L.J.M., D.A.I. Munindradasa & P. Fernando (2012a). A new species of Polypedates Tschudi (Amphibia, Anura, Rhacophoridae) from Sri Lanka. Zootaxa 3498: 63–80; http://dx.doi.org/10.11646/zootaxa.3620.1.5
Wickramasinghe, L.J.M., D.R. Vidanapathirana & N. Wickramasinghe (2012b). Back from the dead: The world’s rarest toad Adenomus kandianus rediscovered in Sri Lanka. Zootaxa 3347: 63–68.
Wickramasinghe, L.J.M., D.R. Vidanapathirana, M.D.G. Rajeev, S.C. Ariyarathne, A.W.A. Chanaka, L.L.D. Priyantha, I.N. Bandara & N. Wickramasinghe (2013a). Eight new species of Pseudophilautus (Amphibia, Anura, Rhacophoridae) from Sripada World Heritage Site (Peak Wilderness), a local amphibian hotspot in Sri Lanka. Journal of Threatened Taxa 5(4): 3789–3920; http://dx.doi.org/10.11609/JoTT.o3099.3789-920
Wickramasinghe, L.J.M., D.R. Vidanapathirana, S. Ariyarathne, G. Rajeev, A. Chanaka, J. Pastorini, G. Chathuranga & N. Wickramasinghe (2013b). Lost and found: one of the world’s most elusive amphibian Pseudophilautus stellatus (Kelaart 1853) rediscovered. Zootaxa 3620 (1): 112–128; http://dx.doi.org/10.11646/zootaxa.3620.1.5
Wickramasinghe, L.J.M., D.R. Vidanapathirana, M.D.G. Rajeev & N. Wickramasinghe (2013c). Rediscovery of, Pseudophilautus hypomelas (Günther, 1876) (Amphibia, Anura, Rhacophoridae) from the Peak Wilderness, Sri Lanka a species thought to be extinct! Journal of Threatened Taxa 5(17): 1–13; http://dx.doi.org/10.11609/JoTT.o3547.5181-93
Appendix 1. Material examined
Pseudophilautus abundus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 3006; Labugama Forest Reserve, Labugama, alt. 78m (06051’N & 080010’E); Paratypes: WHT 2302; 2303; 3457; 3459; 3494; 3496, Dediyagala Forest Reserve, Akuressa, alt. 150m (06010’N & 080026’E); WHT 1711; 3455; 3456; 3495, Kanneliya (Galle), alt. 150m (06015’N & 080020’E).
Pseudophilautus alto (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 2721; Paratypes: WHT 2723; 2718; 2719; 2720; 2722; 2724; Horton Plains National Park, alt. 2135m (06046’N & 080047’E).
Pseudophilautus asankai (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 3507; Paratypes: WHT 3504; 3505; 3506; 5425, Agra Arboretum, Torrington Estate, near Agarapatana, alt. 1665m (06050’36” N, 80040’40” E); WHT 2100, Moray Estate, Rajamally, alt. 1370m (06048’N & 80031’E); WHT 5472, Dayagama Estate, 3rd division, Dayagama, alt. 1830m (06050’N & 80040’E).
Pseudophilautus auratus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 2356; Paratypes: WHT 2357; 2375; 2433; 2782, Morningside Forest Reserve (near Rakwana), alt. 1060m (06024’N & 080038’E); WHT 3282, Handapan Ella Plains (near Rakwana), alt. 1270m (06026’N & 080036’E); WHT 3298, Sinharaja World Heritage Site (near Kudawa), alt. 513m (06025’N & 080025’E).
Pseudophilautus bambaradeniyai Wickramasinghe, Vidanapathirana, Rajeev, Ariyarathne, Chanaka, Priyantha, Bandara & Wickramasinghe 2013. Holotype: NMSL 2013.01.01 NH; Paratypes: DWC 2013.01.001, DWC 2013.01.002, Sripada World Heritage Site, (Adam’s peak), Ratnapura District, alt. 1334m (06048’31.38» N, 080028’14.46» E).
Pseudophilautus caeruleus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 2514; Paratypes: WHT 2511; 2512; 2513, Bogawanthalawa–Balangoda road (near Udugama), alt. 810m (06044’N & 80041’E); WHT 2101A; 2101B, Moray Estate, Rajamally, alt. 1370m (06048’N & 80031’E).
Pseudophilautus cavirostris (Günther 1869). WHT 1294, Haycock (Hiniduma, Galle), alt. 660m (06020’N & 80018’E); WHT 3299; 3300, Sinharaja World Heritage Site, Weddagala, alt. 513m (06025’N & 80025’E); WHT 3389, Kitulgala, alt. 200m (07000’N & 80024’E); WHT 2318; Kosmulla near Neluwa, alt. 450m (06023’N & 80023’E); WHT 2046; 2425, Pathanegala (Knuckles), alt. 1,000m (07033’N & 80044’E); WHT 2045; Kadugannawa, alt. 450m (07015’N & 80030’E); WHT 3483; Pussellawe, alt. 986m (07000’N & 080054’E).
Pseudophilautus cuspis (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 1177(e); Paratypes: WHT 1177(a); 1177(b); 1177(c); 1177(d); 1177(f); 1177(g); 1177(h); 1177(i); 1177(j), Koskulana (near Panapola), alt. 460m (06025’N & 080027’E).
Pseudophilautus dayawansai Wickramasinghe, Vidanapathirana, Rajeev, Ariyarathne, Chanaka, Priyantha, Bandara & Wickramasinghe 2013. Holotype: NMSL 2013.02.01 NH; Paratypes: DWC 2013.01.003, DWC 2013.01.004, Sripada World Heritage Site, (Adam’s peak), Ratnapura District, alt. 1679m (06048’30.89» N 080029’19.18» E).
Pseudophilautus decoris (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: mature female, 23.9 mm SVL, WHT 2358; Paratypes: WHT 3194; 3257; 3258; 3265; 3266; 3267; 3268; 3269; 3271; 3270; 3272; Morningside Forest Reserve (near Rakwana), alt. 1060m (06024’N & 080038’E).
Pseudophilautus dimbullae (Shreve, 1940). Holotype: MCZ A- 20878; Queenwood Estate, Dimbulla.
Pseudophilautus eximius (Shreve, 1940). Holotype: MCZ A-20879, Queenwood Estate, Dimbulla.
Pseudophilautus femoralis (Günther 1864). WHT2701; 2702; 2703; 2704; 2705; 2706; 2707; 3540; 3537; 3538; 3539; 2478; 2479, 7 Jun. 1999, Horton Plains National Park, alt. 2135m (06046’N & 80047’E).
Pseudophilautus fergusonianus (Ahl 1927). WHT 731; 2233, Kumaradola, Monaragala, alt. 305m (06053’N & 081022’E); WHT 1012, Kitulhela (near Moragahapitiya), Monaragala, alt. 520m (07020’N & 81028’E); WHT 2400; 2401; Sera Ella (near Pottotawela), Knuckles, alt. 460m (07035’N & 80045’E); WHT 2038; 2039; 2040, Puwakpitiya (near Laggala), Knuckles, alt. 450m (07034’N & 80045’E); WHT 3360, Deniyaya, alt. 460m (06021’N & 080034’E); WHT 3361; 3362; 3363; 3364; 3365, Pitadeniya (near Watugala), alt. 320m (06022’N & 080028’E) WHT 3168; 3177; 3195, Gannoruwa Forest reserve, Kandy, alt. 684m (07017’N & 080035’E); WHT 3178; 3179; 27; 3180; 3181; 3182, Puwakpitiya, Knuckles, alt. 414m (07034’N & 080044’E); WHT 3229, Hantana, Kandy, alt. 600m (07015’N & 080037’E); WHT 3380; WHT 3381; NMSL 2006.64.1, Medapitiya, Wasgamuwa.
Pseudophilautus folicola (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: mature female, 29.4mm SVL, WHT 2645; Paratypes: WHT 2646; 2647; 2649; 2650; 2651; 2652, Kottawa (Galle), alt. 60m (06006’N & 080020’E).
Pseudophilautus frankenbergi (Meegaskumbura & Manamendra-Arachchi 2005). Holotype: WHT 2554; Paratypes: WHT 2551; 2552; 2555; 2556, Namunukula Peak, alt. 1980m (06056’N & 081007’E); WHT 2726; 2727; 2728; Horton Plains National Park, alt. 2135m (06046’N & 081007’E).
Pseudophilautus fulvus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 2949; Paratypes: WHT 2948, Laggala (Knuckles), alt. 1220m (07033’N & 080044’E); WHT 3112; 3114; 3463, Moussakanda, Gammaduwa, Knuckles, alt. 915m (07034’N & 080042’E); WHT 3121; Puwakpitiya (near Laggala), Knuckles, alt. 450m (07034’N & 080045’E).
Pseudophilautus hallidayi (Meegaskumbura & Manamendra-Arachchi 2005). Holotype: WHT 3575; Paratypes; WHT 3573; 3576; 3577, Hanthana range, Kandy, alt. 510 to 800m (07015’N & 080034’E); WHT 6072, Tonacombe Estate, Namunukula, alt. 1320m (06052’N & 081007’E).
Pseudophilautus hoffmanni (Meegaskumbura & Manamendra-Arachchi 2005). Holotype: WHT 6120; Paratypes: WHT 3222; 3223; 3542, Corbett’s Gap (Knuckles Hills), alt. 1245m (07022’N & 80051’E).
Pseudophilautus hoipolloi (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 2673; Paratypes: WHT 2674; 2675, Haycock (Hiniduma), alt. 660m (06020’N & 080018’E); WHT 2405; 2406; 2407; 2408; 2409, Kahaduwatta (Galle), alt. 15m (06004’N & 080012’E); WHT 3196; 3197; 3198; 3199; 3200, Udugama (Galle), alt. 30m (06014’N & 080020’E); WHT 3201, Kodagoda (Galle), alt. 45m (06002’N & 080023’E).
Pseudophilautus jagathgunawardanai Wickramasinghe, Vidanapathirana, Rajeev, Ariyarathne, Chanaka, Priyantha, Bandara & Wickramasinghe 2013. Holotype: NMSL 2013.03.01 NH; Paratypes: DWC 2013.01.005, DWC 2013.01.006, Sripada World Heritage Site, (Adam’s peak), Ratnapura District, alt. 1642m (06048’31.45” N 080029’10.69” E).
Pseudophilautus karunarathnai Wickramasinghe, Vidanapathirana, Rajeev, Ariyarathne, Chanaka, Priyantha, Bandara & Wickramasinghe 2013. Holotype: NMSL 2013.04.01 NH; Paratypes: DWC 2013.01.007, DWC 2013.01.008, Sripada World Heritage Site, (Adam’s peak), Ratnapura District, alt. 1334m (06048’31.38” N 080028’14.46” E).
Pseudophilautus limbus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 2700; Haycock, alt. 560m (06020’N & 080018’E).
Pseudophilautus lunatus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 3283; Handapan Ella Plains, alt. 1270m (06026’N & 080036’E).
Pseudophilautus macropus (Günther 1869). WHT 3390; 3391; 3392; 3393; 3394; 3395; 3396; 3397; 3398; 3399; 3400; 3401; 3402; 3403; 3404; 3405; 3406, Mousakanda (Gammaduwa), Knuckles, alt. 760m (07034’N & 80042’E); WHT 3183, Divulgahapathana, Knuckles, alt. 603m (07033’N & 80043’E); WHT 5900, Bambaraella, Knuckles, alt. 1260m (07024’N & 80047’E).
Pseudophilautus microtympanum (Günther 1859). WHT 2710; 2708; 2712; 2713; 3436; 3437; 3438; 3439; 3440; 3441; 3442; 3443; 3444; 3446, Horton Plains National Park, alt. 2135m (06046’N & 80047’E); WHT 3260, Agra Arboretum, near Agarapatana, alt. 1555m (06051’N & 80041’E).
Pseudophilautus mittermeieri (Meegaskumbura & Manamendra-Arachchi 2005). Holotype: WHT 3522; Paratypes: WHT 3523; 3524; 3525; 3526, Kottawa, Galle, alt. 60m (06006’N & 080020’E); WHT 2668, Beraliya forest, Elpitiya, alt. 150m (06016’N & 080011’E).
Pseudophilautus mooreorum (Meegaskumbura & Manamendra-Arachchi 2005). Holotype: WHT 5862; Paratypes: WHT 5868; 5869, Hunnasgiriya (Knuckles), alt. 1100m (07023’N & 080041’E); WHT 2477; 3209; 6124, Corbett’s Gap (Knuckles), 1245m (07022’N & 080051’E).
Pseudophilautus nemus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 1319; Haycock (Hiniduma), alt. 660m (06020’N & 080018’E).
Pseudophilautus newtonjayawardanei Wickramasinghe, Vidanapathirana, Rajeev, Ariyarathne, Chanaka, Priyantha, Bandara & Wickramasinghe 2013. Holotype: NMSL 2013.05.01 NH; Sripada World Heritage Site, (Adam’s peak), Ratnapura District, alt. 1559m (06048’36.10» N 080028’59.36» E).
Pseudophilautus ocularis (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 2360; Paratypes: WHT 3273; 2376; 2377; 2378, Morningside Forest Reserve (near Rakwana), alt. 1060m (06024’N & 080038’E); WHT 3288; 3289; 3290; 3291; 3292; 3293, Handapan Ella Plains (near Suriyakanda), alt. 1270m (06026’N & 080036’E).
Pseudophilautus papillosus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 3284; Handapan Ella Plains (near Rakwana), alt. 1270m (06026’N & 080036’E).
Pseudophilautus pleurotaenia (Boulenger 1904). WHT 3176; 5824; Gannoruwa Forest reserve, Kandy, alt. 684m (07017’N & 080035’E); WHT 5860; 5861; Nawalapitiya, alt. 700m (07003’N & 080032’E).
Pseudophilautus poppiae (Meegaskumbura & Manamendra-Arachchi 2005). Holotype: WHT 3285; Handapan Ella Plains (near Suriyakanda), alt. 1270m (06026’N & 080036’E); Paratypes: WHT 2030; 2029; 2475; 2778; 2781; 3533; 3534; 3535; ; 3536, Morningside (near Rakwana), alt. 1060m (06024’N & 080038’E).
Pseudophilautus procax (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 3279; Paratypes: WHT 2786; 2787; 3277; 3278; 3280; 3281, Morningside Forest Reserve (near Rakwana), alt. 1060m (06024’N & 080038’E).
Pseudophilautus puranappu Wickramasinghe, Vidanapathirana, Rajeev, Ariyarathne, Chanaka, Priyantha, Bandara & Wickramasinghe 2013. Holotype: NMSL 2013.06.01 NH; Paratypes: DWC 2013.01.009, DWC 2013.01.010, Sripada World Heritage Site, (Adam’s peak), Nuwara Eliya District, alt. 2106m (060 48’26.03” N 080029’58.09”E).
Pseudophilautus regius (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 3122; Paratypes: WHT 5827; 5828; 5829, Angammadilla, Polonnaruwa, alt. 90m (07051’N & 080055’E).
Pseudophilautus reticulatus (Günther 1864). WHT 2286; 2287; 2290; 2291, Kosmulla (near Nelluwa), alt. 320m (06024’N & 80023’E); WHT 2288; 2289, Dediyagala Forest Reserve, Akuressa, alt. 150m (06010’N & 80026’E); WHT 2344, Yagirala, alt. 30m (06022’N & 80010’E); WHT 3230, Induruwa, (Ratnapura), alt. 150m (06045’N & 80026’E); NMSL 2006.66.1; Gileemale forest, alt. 150m (06045’N & 080026’E); WHT 2520; 2521, Check Poleat Gap (near Norton Bridge), alt. 800m (06056’N & 80030’E); WHT 3366, Haycock (Hiniduma), alt. 660m (06020’N & 80018’E)
Pseudophilautus rus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 3474; Paratypes: WHT 3475; 3476; WHT 3477, Kiribatkumbura (near Peradeniya), alt. 450m (07016’N & 080034’E); WHT 5437; 5439; 5436; WHT 5440; 5438; 5432; 5435; 5434, Pilimatalawa (near Peradeniya), alt. 658m (07015’N & 080034’E).
Pseudophilautus samarakoon Wickramasinghe, Vidanapathirana, Rajeev, Ariyarathne, Chanaka, Priyantha, Bandara & Wickramasinghe 2013. Holotype: NMSL 2013.07.01 NH; Paratypes: DWC 2013.01.011, DWC 2013.01.012, Sripada World Heritage Site, (Adam’s peak), Ratnapura District, alt. 1334m (06048’31.38” N 080028’14.46” E).
Pseudophilautus sarasinorum (Müller 1887). WHT 2480; 2481; 2482; 2483, Bogawanthalawa-Balangoda road, alt. 1300m (06045’N & 080042’E); WHT 2426; 2427; 2428; 2429, Corbett’s Gap, alt. 1000m (07022’N & 080050’E).
Pseudophilautus schmarda (Kelaart 1854). Neotype: WHT 3353; Horton Plains National Park, alt. 2135m (06046’N & 80047’E).
Pseudophilautus silus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 3453; Paratypes: WHT 3451; WHT 3452; WHT 3454; WHT 3460, Agra Arboretum, near Agarapatana, alt. 1555m (06051’N & 080041’E); WHT 3412, Tangamalai Sanctuary, near Haputale, alt. 1600m (06046’N & 080055’E);
Pseudophilautus silvaticus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 3295; Paratypes: WHT 3296, 3378; 3379; 3461; 3462, Handapan Ella Plains (near Suriyakanda), alt. 1270m (06026’N & 080036’E); WHT 3275; WHT 3276, Morningside Forest Reserve (near Rakwana), alt. 1060m (06024’N & 080038’E); WHT 3310; 3316, Sinharaja World Heritage Site (near Kudawa), alt. 513m (06025’N & 080025’E).
Pseudophilautus simba (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 3464; Paratypes: WHT 3465; WHT 3466; WHT 3467, Morningside Forest Reserve (near Rakwana), alt. 1060m (06024’N & 080038’E).
Pseudophilautus sirilwijesundarai Wickramasinghe, Vidanapathirana, Rajeev, Ariyarathne, Chanaka, Priyantha, Bandara & Wickramasinghe 2013. Holotype: NMSL 2013.08.01 NH; Paratypes: DWC 2013.01.013, Sripada World Heritage Site, (Adam’s peak), Ratnapura District, alt. 1679m (06048’30.89” N 080029’19.18” E).
Pseudophilautus sordidus (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 2379; Paratypes: WHT 2383; 2384; 2385; 2386; 2389; 2390; 2391; 2393; 2395; 2397; 2398, Kanneliya Forest Reserve (Galle), alt. 150m (06015’N & 080020’E); WHT 2380; WHT 2381, Haycock (Hiniduma), alt. 660m (06020’N & 080018’E); WHT 2382, Haycock (Hiniduma), alt. 150m (06018’N & 080019’E); 2387; WHT 2396, Millawa Forest Reserve (near Morawaka), alt. 150m (06017’N & 080028’E); WHT 2988, Welikanna, Waga (near Labugama), alt. 78m (06051’N & 080009’E); WHT 2998, Labugama Forest Reserve, alt. 78m (06051’N & 080010’E); WHT 3303; 3304; 3306, Sinharaja Forest Reserve (Halmandiya), alt. 513m (06025’N & 080025’E); NMSL 2006.65.1. Denenakanda, Peak Wilderness.
Pseudophilautus steineri (Meegaskumbura & Manamendra-Arachchi 2005). Holotype: WHT 3210; Paratypes: WHT 3519; 3521; 3520, 6116; 3518, Corbett’s Gap (Knuckles Hills), alt. 1245m (07022’N & 080051’E).
Pseudophilautus stictomerus (Günther 1876). WHT 1173; 2402, Kottawa (Galle), alt. 60m (06006’N & 080020’E); WHT 2403; 2404, Kanneliya (Galle), alt. 150m (06015’N & 080020’E); WHT 3301, Sinharaja Forest (near Kudawa), alt. 513m (06025’N & 080025’E); WHT 3355; 3356; 3357; 3358; 3359, Kosmulla (near Nelluwa), alt. 320m (06024’N & 080023’E).
Pseudophilautus stuarti (Meegaskumbura & Manamendra-Arachchi 2005). Holotype: WHT 3208; Paratypes: WHT 3207; 3206; 3218, 3527, 357 Corbett’s Gap (Knuckles Hills), alt. 1245m (07022’N & 080051’E).
Pseudophilautus viridis (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 5127; Paratypes: WHT 3488; 3489; 5104; 5105; 5106; 5099; 5100; 5103; 5126, Agra Arboretum, near Agarapatana, alt. 1555m (06051’N & 080041’E); WHT 2763; 2764; 2765; 2766; 2767; 2768, 4999, Ambewela (near radio station), alt 1830m (06053’N & 080048’E).
Pseudophilautus zorro (Manamendra-Arachchi & Pethiyagoda 2005). Holotype: WHT 3169; Paratypes: WHT 3175; 3204; 3205, Gannoruwa Forest Reserve, Kandy, alt. 684m (07017’N & 080035’E); WHT 3508, Tiverton Estate (near Peradeniya), alt. 450m (07016’N & 080034’E).