Journal of Threatened Taxa | www.threatenedtaxa.org | 26 May 2016 | 8(5): 8745–8755

Diet of the Four-horned Antelope Tetracerus quadricornis (De Blainville, 1816) in the Churia Hills of Nepal
Amar Kunwar 1, Raju Gaire 2, Krishna Prasad Pokharel 3, Suraj Baral 4 & Tej Bahadur Thapa 5
1,4,5 Central Department of Zoology, Tribhuvan University, TU Rd, Kirtipur 44618, Nepal
2 Kathmandu Model College, Bagbazar GPO Box No. 4470. Kathmandu Nepal
3 Wildlife Ecology and Management, University of Freiburg, Tennenbacher Str.4, D - 79106 Freiburg, Germany
1 amar.kwr@gmail.com (corresponding author), 2 uniqueraj2010@yahoo.com, 3 kp.pokharel@gmail.com,
4 baral.hector@gmail.com, 5 tbthapa@cdztu.edu.np
doi: http://dx.doi.org/10.11609/jott.1818.8.5.8745-8755
Editor: Mewa Singh, University of Mysore, Mysuru, Kerala. Date of publication: 26 May 2016 (online & print)
Manuscript details: Ms # 1818 | Received 07 February 2015 | Final received 03 May 2016 | Finally accepted 06 May 2016
Citation: Kunwar, A., R. Gaire, K.P. Pokharel, S. Baral & T.B. Thapa (2016). Diet of the Four-horned Antelope Tetracerus quadricornis (De Blainville, 1816) in the Churia Hills of Nepal. Journal of Threatened Taxa 8(5): 8745–8755; http://dx.doi.org/10.11609/jott.1818.8.5.8745-8755
Copyright: © Kunwar et al. 2016. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: Self-funded.
Conflict of Interest: The authors declare no competing interests.
Author Details: Mr. Amar Kunwar has completed his master’s degree in Zoology, specialization in wildlife ecology and environment from Central Department of Zoology. Currently, he is working in ecological conservation of Blackbuck Antelope cervicapra and threatened small mammals of Nepal. He is interested in ecological conservation and research of prey-species of Nepal, especially Ungulates. Mr. Raju Gaire has completed his master’s degree in Zoology, specialization in Ecology from Central Department of Zoology and conducting researches on ecological studies of Barking deers Muntiacus vaginalis in Nepal. He is interested in research of prey-species. Dr. Krishna Prasad Pokharel has completed his PhD in Niche Differentiation of Solitary Ungulates in Lowland Nepal and currently is a scientific staff at Chair of Wildlife Ecology and Management, University of Freiburg, Germany. Mr. Suraj Baral has completed his master’s degree in Zoology from Central Department of Zoology in Ecology and conducting ecological researches on Herpetofauna of Nepal. Dr. Tej B. Thapa is a professor at Central Department of Zoology, Tribhuvan University, Nepal. He has completed his PhD in habitat suitability evaluation for Leopard Panthera Pardus using remote sensing and GIS. His area of interests are big cat ecology and predator-prey relationships.
Author Contribution: AK - surveyed in the field, conducted laboratory work, calculated, analyzed and prepared the manuscript. RG - surveyed in the field, collected samples and did laboratory work. KPP - guided in field methods and collected samples in the field. SB - analysed the data and worked in laboratory. TBT - guided in sample collection, laboratory work, identification of plant species and analysis and supervised the whole work.
Acknowledgements: We are thankful to the Central Department of Zoology, Tribhuvan University for providing laboratory support and Department of National Parks and Wildlife Conservation and National Trust for Nature Conservation, Nepal for field support. We owe special thanks to Dr. Narendra M.B. Pradhan for providing references and microphotographs of some plants. we are grateful to Mr. Buddi Sagar poudel for providing important laboratory advices.
Abstract: The food composition of the Four-horned Antelope Tetracerus quadricornis was studied in the Churia Hills of Nepal during summer, monsoon and the winter seasons of 2012–2013. Microhistological technique was used to determine the diet. The Four-horned Antelope was found to be a mixed feeder feeding on trees, shrubs, forbs, grasses and climbers. Trees and shrubs contribute the major percentage of diet in all the three seasons. The Gramineae family is consumed in highest proportion. Mitragyna parvifolia, Bridelia retusa, Bambusa vulgaris, Hymenodictyon sp. and Ziziphus mauritiana are major tree species while Barleria cristata, Pogostemon benghalensis, Achyranthes sp., Clerodendrum viscosum are among shrubs. Ageratum conyzoides and Blumea virens are the main forbs Eulaliopsis binata and Imperata cylindrica are the principal grass species. Climber Trachelospermum lucidum is consumed in a small proportion. Grasses in monsoon were consumed distinctly at a higher percentage than during the other two seasons. The Four-horned Antelopes are concentrated feeders and browsers with a generalized feeding strategy. Similar studies need to be conducted in other landscapes and with sympatric and potential competitor species to understand its niche overlaps and degree of competition.
Keywords: Bardia National Park, Browser, food composition, generalist, Gramineae, micro-histological analysis, mixed feeder, niche breadth.
INTRODUCTION
Quantifying diet is an important aspect of studying animal ecology. Without full understanding of what animals eat, why they eat, where they eat and variations in feeding over time, species specific management is nearly impossible. Efficient management of ungulates requires a detailed knowledge of their diets and food habits (Holechek et al. 1982; Mofareh et al. 1997; Sandoval et al. 2005). Knowing the diet composition of herbivores is important for understanding their foraging ecology and for mediating the effects they have on vegetation and ecosystems (Barcia et al. 2007). The quality of food significantly influences reproduction, growth and survival of animals (Pekins et al. 1998) and consequently on their population dynamics. The dietary studies are more important for patchily distributed (Sharma et al. 2013), threatened and low density species (Sharma et al. 2009) like the Four-horned Antelope Tetracerus quadricornis (de Blainville 1816).
The Four-horned Antelope (F-hA), endemic to India and Nepal, is one of the unique tropical antelopes and smallest Asian bovid with four horns in the male. It chooses open habitats (Prater 1971; Krishna et al. 2009) and also prefers forested areas to open grasslands (Sharma & Rahmani 2005), with an undulating or hilly terrain (Prater 1971; Sankhala 1977). Historically, the F-hA was distributed from the Terai region of Nepal in the north, to the Nilgiri Hills in the south and Bengal region in the east in India to Sind province in the west (Blanford 1888; Jerdon 1874; Murray 1884), but are currently extirpated from Pakistan (Roberts 1997). Presently, they are reported from four protected areas of Nepal; Bardia National Park (Karki 1998; Steinheim et al. 2005; Pokharel 2010), Chitwan National Park (Gurung and Singh 1996, Pokharel 2012), Parsa Wildlife Reserve (Shrestha 2001) and Banke National Park (DNPWC 2010) and in all states of India except Kerala (Rice 1990). Although, this Vulnerable species (IUCN 2013) occurs only in Nepal and India (Rahmani 2001), baseline information about their distribution, feeding ecology, habitat requirements, population density and present status from Nepal are lacking.
Very little information on the food habits of the F-hA is available from India, through cafeteria experiments (Berwick 1974; Sharma et al. 2009), direct observations (Sharma et al. 2005) and micro histological analysis (Baskaran et al. 2011). The species is a browser (Pokharel et al. 2015) and forages selectively on nutritious plant parts such as fruits, flowers and fresh leaves (Berwick 1974; Sharma et al. 2005; Baskaran et al. 2011). No studies on its feeding habits have been reported from Nepal.
This paper aims to assess the diet composition of the F-hA at the plant category and plant species level in Nepal. Field samplings were done in three different seasons during summer (March–April 2012), monsoon (September 2012) and winter (January 2013).
MATERIALS AND METHODS
Study Area
The study was conducted in the Churia Hills of the northern slopes of 968km2 Bardia National Park (28015’–28040’N & 81015’–81040’E), located in western Tarai and Siwalik of Nepal (Fig. 1). The altitudinal gradient ranges from 152m at the south-western corner of the park to an elevation of 1,441m at Sukurmala to the crest of Churia ridge (Dinerstein 1979). The park can be classified into five distinct land types; the Churia (Siwaliks), the Bhabar foothills, the alluvial flat lands, the riverine floodplains and the Babai Valley. The park experiences sub-tropical monsoon climate with three distinct seasons: monsoon, winter and summer. The park holds two major eco-regions, namely the Terai-duar savannas and grasslands, and the Himalayan sub-tropical broadleaved forests. The vegetation is sub-tropical, consisting of a mosaic of early successional floodplain communities along the Babai River and its tributaries, and with large areas of climax Shorea robusta forest on the upper drier land. Around 76% of the total park area is covered by forest; 52% of the total plant species are trees, 20% are shrubs while the remaining 8% are herbs (Bhuju et al. 2006). The major vegetation types are Sal forest, riverine forest, mixed hardwood forest, wooded grasslands, phantas and tall alluvial flood plain grassland. Important flora in the park includes Shorea robusta, Terminalia tomentosa, Mallotus philippensis, Acacia catechu, Dalbergia sissoo, Bombax ceiba, Pinus roxburghii, Buchanania latifolia, Dillenia pentagyna, Murraya koenigii, Colebrookea oppositifolia, Pogostemon benghalensis, Imperata cylindrica, Saccharum sp. (Shrestha et al. 1997). It harbors 59 species of mammals, 407 species of birds, 42 species of reptiles and amphibians, and 124 species of fishes (BPP 1995; DNPWC 2001).
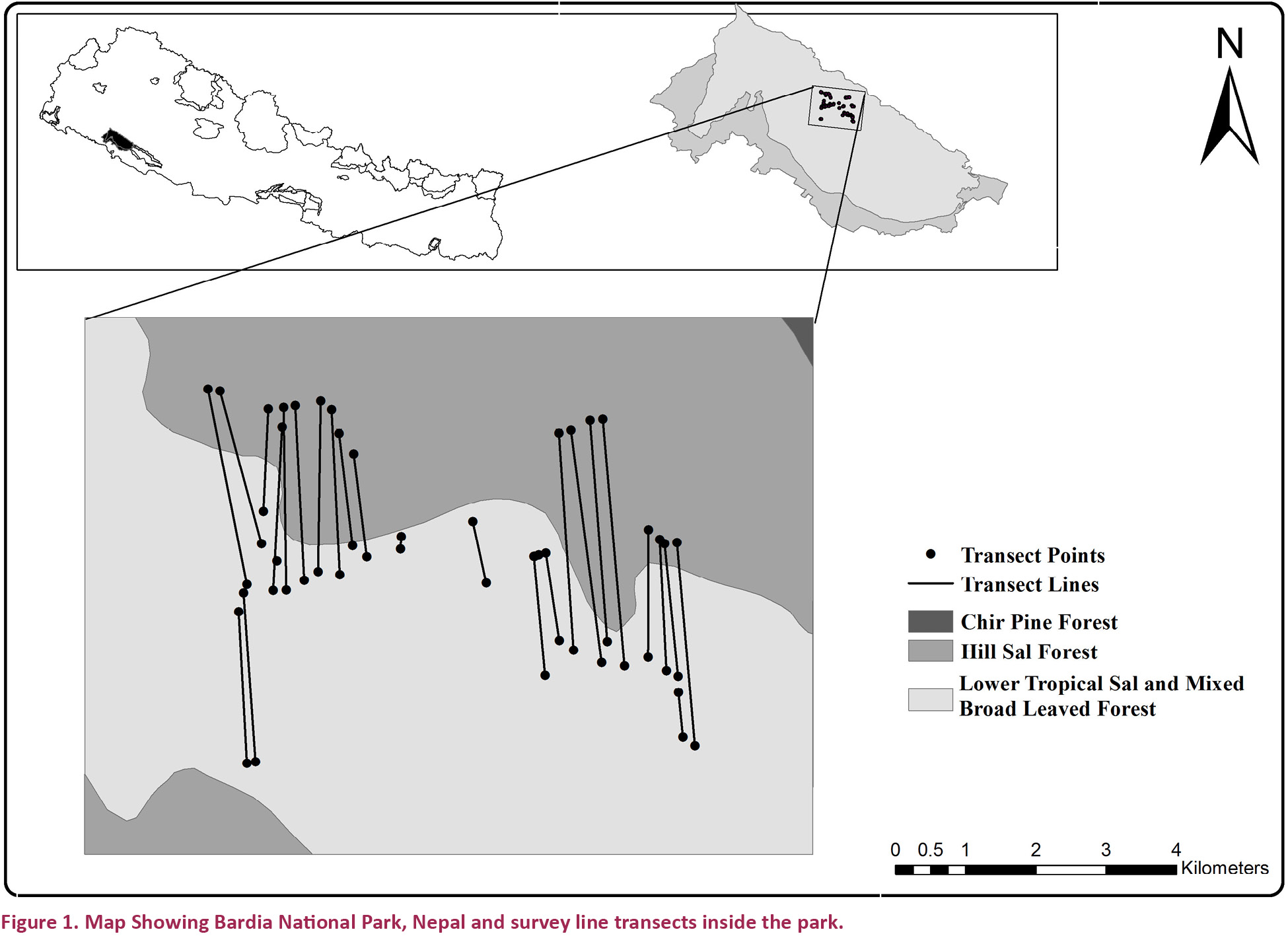
Dietary analysis
The diets of the F-hA were determined following a standard micro-histological technique (Norbury 1988). The epidermal features of plants in the F-hA faeces were identified with the help of reference slides prepared from 104 plant species collected from the study area. The reference plant samples were dried in an oven at 600C and the dried samples were separately ground with an electric blender and sieved in a mesh size of 1–0.3 mm. The powder retained on the 0.3mm sieve was chosen as the final sample for slide preparation. The final sample was placed in Petri dishes and bleached with 4% sodium hypochlorite for 6–24 hours at room temperature to remove mesophyll tissue and to render the epidermis identifiable. The bleached contents were then rinsed well in a sieve and then treated with a few drops of staining substance-gentian violet solution (1g/100m water) for 10 seconds and again rinsed. The stained fragments were mounted on standard microscope slides in a glycerin medium and covered with a cover slip. A similar process was followed for the faecal samples. Both reference slides and faecal slides were observed at different magnifications; 100x, 200x and 400x with a compound microscope and each fragment was photographed using a digital camera for microscope (DCM510; USB2.0; 5M pixel, CMOS chip) in a laptop using software- ScopeTek Scope Photo; Version: x64, 3.1.615 (http://www.scopetek.com).
For each fecal sample, non-overlapping and distinguishable 30 fragments, observed while moving the slides from left to right in the microscope, were identified considering the specific histological feature of the epidermis, i.e., epidermal cell shape, size and arrangement; vascular vessels type; stomata type and arrangement; venation characteristics; shape and arrangements of hairs and trichome, crystal types, etc.
To analyze the diet data, four levels of classifications were constructed into which plant fragments were assigned: (1) functional group (F.G.): (i) grasses, (ii) forbs, (iii) shrubs, (iv) climbers and (v) trees; (2) broad taxonomic group (B.T.): (i) monocots and (ii) dicots; (3) family; and (4) species. Those plants, which could not be identified to species or genera levels, were grouped into “unknown grass”, “unknown forbs”, “unknown shrub”, “unknown climber” and “unknown tree”.
The diet composition was expressed as a percentage of occurrence (O%) (Cavalini & Lovari 1991).
Number of fragments of each food
Percentage of occurrence (O %) = ––––––––––––––––––––––––––X 100
Total number of plant fragments read
Chi-square test (χ2) was used to determine the significance of variation in preferences of functional plant categories in three different seasons using program SPSS (SPSS version 16.0.0).
To evaluate the degree of selectivity of plant species in the diet, Levin’s measure of niche breadth (Levins 1968), was used.
The equation is
1
B = n
∑ = 1 pi2
i
Where, pi = Percentage of total samples belonging to species i (i= 1,2,3,……, n)
n = total number of plant species in all samples.
Diversity was standardized to a scale of 0.0 to 1.0 by using Hurlbert’s method (Krebs 1999);
Bs= B - 1 / n - 1
Where, Bs = Levins’s standardized niche breadth,
B = Levins’s measure of niche breadth, and n = number of possible resource states.
To evaluate whether the FHA is a browser or grazer, the monocots and dicots were further assigned as grass and browse respectively (Jarman 1974; Shipley 1999) and their ratio was expressed in terms of percentage.
RESULTS
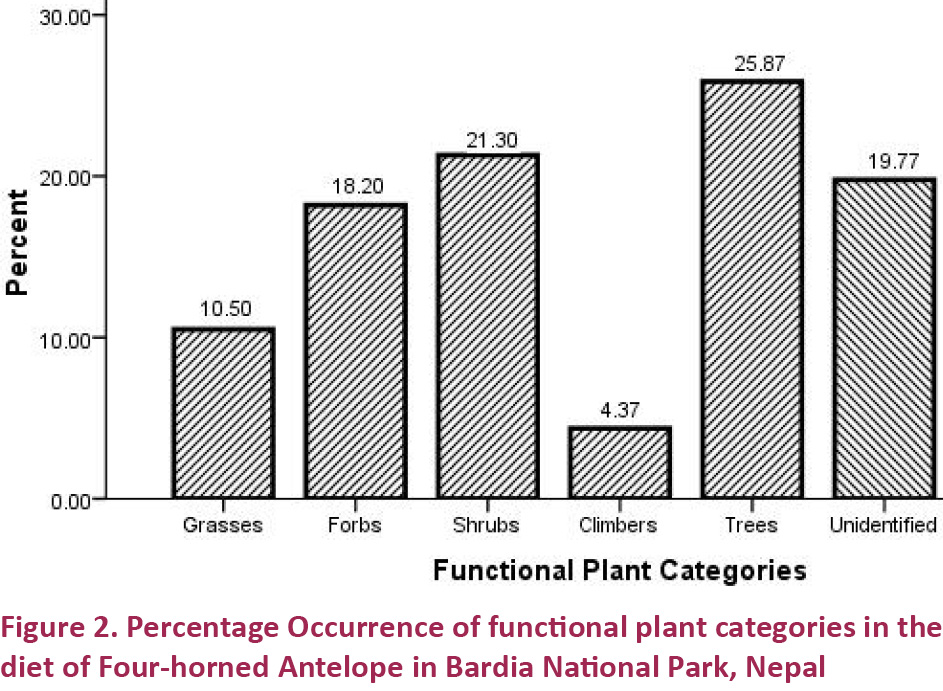
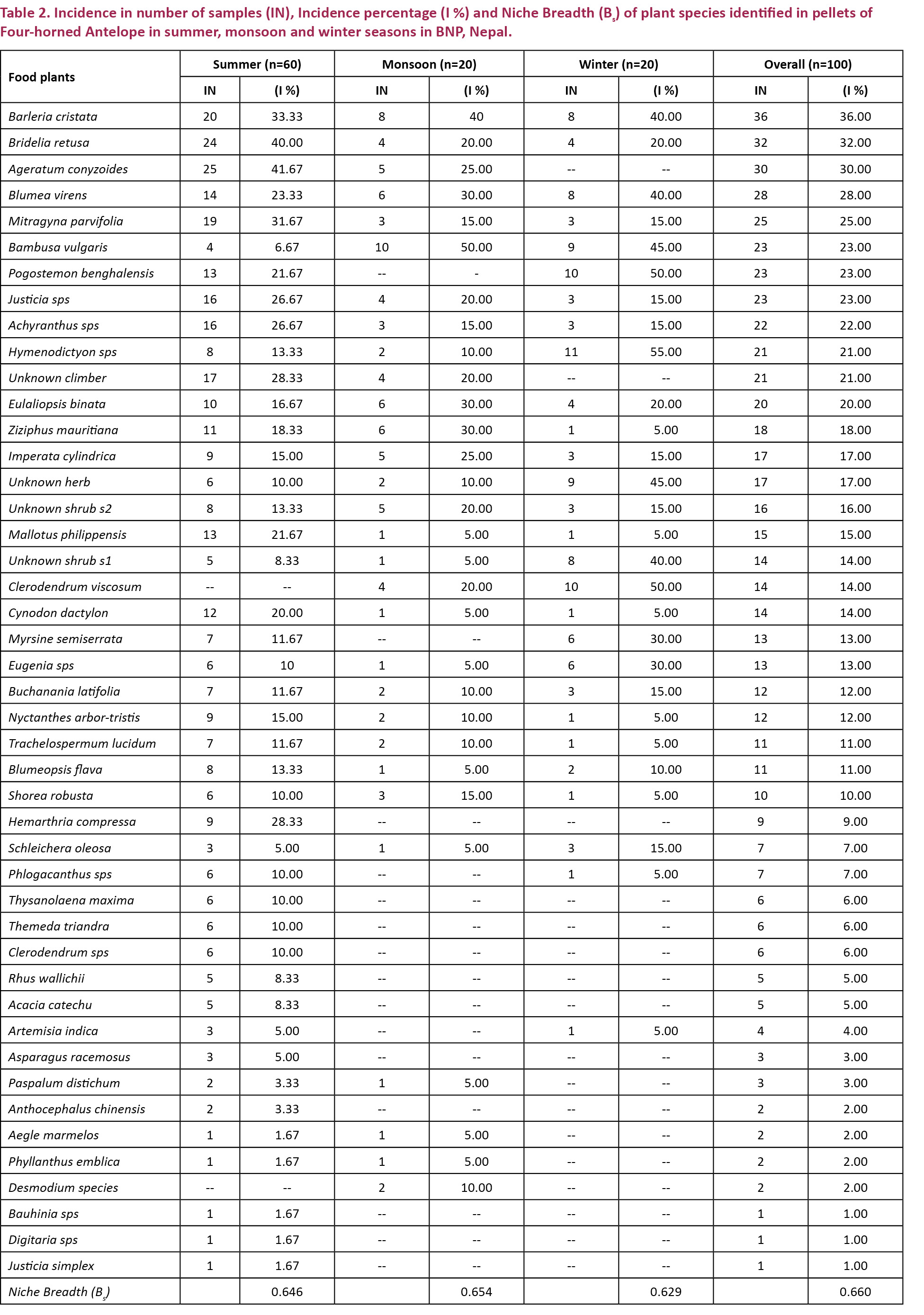
We analyzed 100 pellet samples from summer (n=60), monsoon (n=20) and winter (n=20). Results revealed that a total of 45 species of plants belonging to 20 different families including 15 tree species, 15 shrubs, eight forbs, five grasses and two climbers were part of the diet. Of the total food plant species, 10 were monocots and 35 were dicots (Table 1). Trees contributed the major proportion (25.87%) of the diet followed by shrubs (21.3 %), forbs (18.2 %), and grasses (10.5 %). Climbers contributed to the least proportion of occurrence (4.36%) in the diet of the F-hA (Fig. 2). The F-hA diet was dominated by the browse plant species. The ratio of browse to grass was 78.03% / 21.97 %, showing a strong affinity towards consumption of browse plant species.
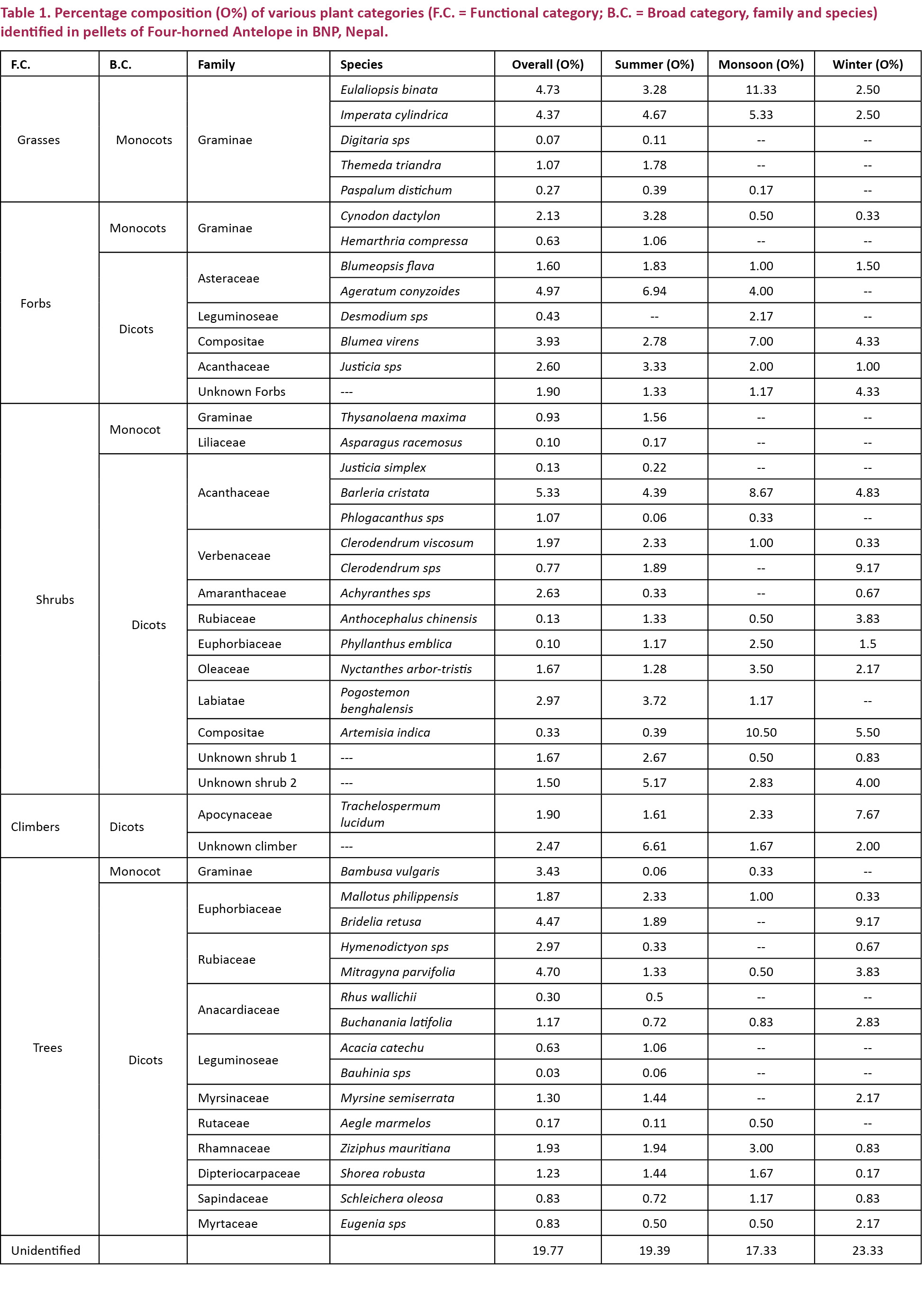
Plant species belonging to the family gramineae were consumed in highest proportion (17.64%) followed by Acanthaceae (9.13%), Rubiaceae (7.8%), Asteraceae (6.56%), Euphorbiaceae (6.4%) and others (Fig. 3).
Mitragyna parvifolia (4.7%), Bridelia retusa (4.47%), Bambusa vulgaris (3.43%), Hymenodictyon orixense (2.97%), Ziziphus mauritiana (1.93%), Mallotus philippensis (1.86%), Buchanania latifolia (1.67%), Myrsine semiserrata (1.30%) and Shorea robusta (1.23%) were the major tree species consumed while Aegle marmelos, Schleichera oleosa, Acacia catechu, Rhus wallichii, Bauhinia sp. and Eugenia sp. had their occurrence below one percent. Likewise, Barleria cristata was the dominant food item among the shrubs with its percentage occurrence of 5.33%, followed by Pogostemon benghalensis (2.96%), Achyranthes (2.63%), Clerodendrum viscosum (1.96%), Nyctanthes arbor-tristis (1.66%) and by Phlogacanthus spp. (1.067%). Other shrubs; Thysanolaena maxima, Asparagus racemosus, Anthocephalus chinensis, Phyllanthus emblica, Artemisia indica, Justicia simplex and Clerodendrum sps were present in small amounts. Forbs were also present in remarkable proportions in the diet. They contributed 18.2% of the overall diet. Ageratum conyzoides (4.97%) and Blumea virens (3.93 %) were the main forbs preferred by the animal. Cynodon dactylon (2.13%), Justicia spp. (2.6%), Blumeopsis flava (1.6%), Hemarthria compressa (0.63%) and Desmodium spp. (0.43%) were other forbs fed. Eulaliopsis binata and Imperata cylindrica were the principal foods among grass species. They contributed 4.73% and 4.36% respectively. The rest, Themeda triandra (1.0%), Paspalum distichum (0.26%) and Digitaria sps (0.06%) were also present in a minority. In addition, two climbers; Trachelospermum lucidum and one unknown climber were used up to 1.9% and 2.46% respectively as food.
Seasonal diet composition
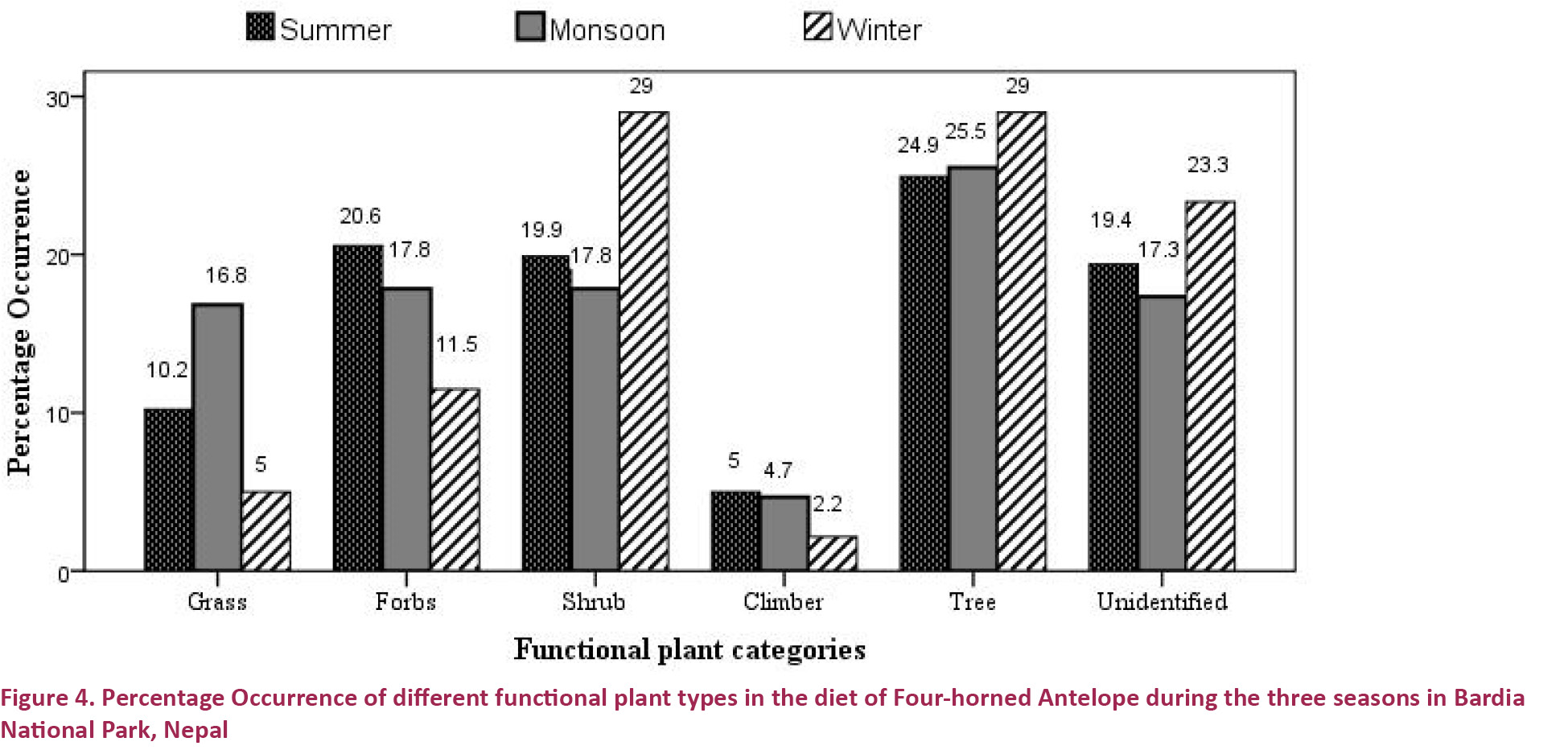
Trees contributed the major percentage of diet in all the three seasons. Shrubs were consumed relatively in a higher proportion in winter (29.00%) than in summer (19.89%) or monsoon (17.83%). The preference order of forbs followed from summer (20.56%) to monsoon (17.83 %) and winter (11.50%). Grasses during the monsoon were consumed distinctly at a higher percentage (16.83 %) than in summer (10.22%) or winter (5.00%). Climbers contributed a small proportion during all the three seasons (Fig. 4). Browse to grass ratio was maximum during the winter at 85.86% / 14.14% while it declined to 80.85% / 19.15% during the summer and to 66.34% / 14.14% during the monsoon.
The frequency of consumption of functional plant categories (grasses, forbs, shrubs, climbers and trees) in the diet of F-hA varied among the seasons (χ2 = 88.38, d.f. = 8, p-value <0.001). F-hA ate more diverse food species in the summer (43 species) than in the monsoon (30 species) or winter (28 species).
Summer season diet
During summer, the F-hA consumed 43 species of plants belonging to 20 families (Table 1). Trees were the major diets consumed (24.95%). Altogether 15 tree species, belonging to 11 families occurred in the summer diet. Forbs with seven species were the second important category contributing 20.56% of the diets. Furthermore, 19.88 percentage of occurrence was lent by shrubs, 10.22% by grass and 5.00% by climbers. The F-hA ate 14 shrubs, five grasses and two climbers during summer (Table 1). In this season, the Four-horned Antelope fed on 32 browse species of plants and 10 grasses (Table 1) and the browse to grass ratio that was obtained was 80.85% / 19.15%.
Monsoon season diet
During the Monsoon season, the F-hA consumed 30 species belonging to 17 families comprising three grasses, seven forbs, seven shrubs, two climbers and 11 trees (Table 1). Trees were consumed at the highest proportion (25.50%) while shrubs and forbs were consumed in equal proportion (17.83 %) during this season. Three species of grass (Graminae) contributed 16.83 percent of occurrence in the diet. Two species of climber were also available in the diet and added 4.67% of occurrence to the total diet (Fig 4). In this season, F-hA fed on 25 dicots (browse) plants and five monocots (grasses) (Table 1) and the browse to grass ratio was found to be 66.34% / 14.14%.
Winter season diet
The F-hA ate 28 species of plants during the winter season belonging to 20 families (Table 1). The F-hA showed equal affinity towards the consumption of trees and shrubs; each contributed 29.0% of the winter diet. Five species of forbs shared their proportion of the diet by 11.5%. Only two species of grass and a climber were consumed in the winter. They contributed 5.0% and 2.17% respectively (Fig. 4). In this season, the F-hA fed on 24 dicot (browse) species of plants and four monocots (grasses) (Table 1) and the browse to grass ratio that was obtained was 85.86% / 14.14%.
Niche breadth
Standardized Levin’s Measure of niche breadth (Bs) of the food plants present in the diet was found to be 0.66 (Table 2) showing that the F-hA utilize broad categories of forage plants and adopt a generalized feeding strategy. This feeding strategy was found in all three seasons; summer (Bs=0.646), monsoon (Bs=0.654), and winter (Bs=0.629) (Table 2).
DISCUSSION
Browse species are the most important food for F-hA, constituting two-third (66.95%) of the overall diet proportion while grass species constituted only 13.68% (the rest, 19.77% remained unidentified). This contradicts the findings of Baskaran et al. (2011), who reported an equal proportion of grass and browse. The browse domination in the diet, in this study, supports the results of the feeding observations made on free ranging (Berwick 1974) and tamed antelopes (Solanki & Naik 1998; Sharma 2006) from central and western India. According to Hofmann (1989), concentrate feeders choose a mixed diet with grasses less than 25% and show a remarkable degree of forage selectivity. The Four-horned Antelopes in the Churia Hills of Bardia National Park are concentrate feeders, consuming different proportions of various plant species and forage categories (grasses, forbs, shrubs, climbers and trees). On the whole, trees constituted the major part of the diet contributing 25.86%, followed by shrubs (21.30%), forbs (18.20%), grasses (10.50%) and climbers (4.37%). The study made by Baskaran et al. (2011) in tropical forests of southern India during dry season revealed grasses as the major constituent of the Four-horned Antelope diet (28.6%) followed by trees (8.0%), shrubs (5.6%) and herbs (6.7%). They argue that during the dry season, grass becomes too coarse and less nutritive hence this antelope depends on both browse and grass, and appears to adapt its feeding according to availability. The shrub Berlaria cristata of the family Acanthaceae is the most preferred plant by this species. The other plants, in decreasing order of preference, are Ageratum conyzoides (Asteraceae), Eulaliopsis binata (Graminae), Mitragyna parvifolia (Rubiaceae), Bridelia retusa (Euphorbiaceae), Imperata cylindrica (Graminae), Blumea virens (Compositae), Bambusa vulgaris (Graminae), Hymenodictyon orixense (Rubiaceae), Pogostemon benghalensis (Labiatae), Achyranthes sp. (Amaranthaceae), Cynodon dactylon (Graminae), etc. (Table 1). The cafeteria experiments of Berwick (1974) in Gir forest ecosystem, India, and Sharma et al. (2009) in Van Vihar National Park cum Zoo in Bhopal, India, showed that Ziziphus mauritiana was the most preferred plant for the tamed F-hA (>24%). This study reveals a contrasting result showing minimum affinity towards the consumption of Z. mauritiana in the wild. Although highly palatable, the thorns of Z. mauritiana inhibit its consumption in the natural habitats (Berwick 1974). The F-hA in the Churia hills of Bardia National Park do not show much preference towards climber plants. Only two climbers (Trachelospermum lucidum and one unknown) were consumed to a small percentage.
Variation in the consumption of different plant types by F-hA in different seasons have been related to changes in the chemical composition of food plants (Field 1971; Sukumar 1989). Increased browsing plants during summer and winter seasons and grazing plants during monsoon are related to protein and fiber content. Plant species differ in protein and fiber contents which influences food choice (Klaus-Hugii et al. 1999) and digestibility of animals (Harbone 1991). The food selectivity in F-hA might be affected by the nutritional requirements, the need to decrease fiber intake, and maximization of protein intake in order to increase digestibility. Although inadequate sample sizes in monsoon and winter seasons make it inappropriate to extrapolate this result to the diet of all the seasons, still, it is rather fair to say that the Four-horned Antelopes inhabiting Churia hills prefer browse species. The other closely related species of this antelopes are also found to be browsers; such as African Bushbuck Tragelaphus scriptus (Odendaal 1983), Kudu Tragelaphus species (Owen-Smith 1993) and Eland Taurotragus oryx (Buys 1990). The F-hA preferred to eat tree species in all three seasons but in different proportions. The F-hA consumed tree species more in the winter season but decreased its consumption during the summer and the monsoon. Forbs were consumed more in summer than in the monsoon or winter. The consumption of grasses was more in monsoon than in summer and winter. The consumption of grass was at a higher proportion during monsoon season, as the Four-horned Antelopes are known to consume grasses more in the monsoon season while they have specialized foraging preferences during the other seasons (Rodger & Panwar 1988). The grasses in monsoon are rich in nutrient content compared to other seasons (Sukumar 1989) while in other seasons they become too coarse and poor in nutrient contents (Baskaran 1998).
Consumption patterns of different food plants vary with seasons. In environments with prominent seasonal changes, food resources are commonly limited and dietary quality and quantity vary highly during dormant seasons. Consequently, the highest intake of digestible nutrients by herbivores occurs during the summer or rainy seasons (Parker et al. 2009). In the Churia Hills of Nepal too, the plant resource heterogeneity, duration of the dormant season, and rate of decline in forage quality all must have affected the seasonal cycle of food intake by the F-hA.
Nearly one fifth of the total plant fragments remained unidentified in this study. In-vitro digestibility greatly influences the results of microhistological analysis particularly in the estimation of grass and forbs content (Vavra & Holechek 1980). The digestive efficiency of deer is very high and so the ingested plant parts are almost degraded (Korschgen 1971). The F-hA prefers fruits, flowers and fresh leaves (Berwick 1974; Sharma et al. 2005; Baskaran et al. 2011) which are highly degradable. Thus, this high percentage of unidentified plants in the diet could be due to high mastication and efficient digestion by the animal. Also, the biases subjected to microhistological analysis, like sample preparation (Vavra & Holechek 1980) poor training of technician (Holechek & Gross 1982) and differential digestibility of diet components (Holechek & Valdez 1985) may have influenced in resulting in the large percentage of fecal plant fragments that remain unclassified.
From the range of food plants eaten, the F-hA in the Churia Hills of Nepal are found to adopt a generalized feeding strategy. This strategy enables them to utilize a mix of dietary food plants to obtain the best source of different important nutrients that an animal requires and detoxify large amounts of chemically similar plant secondary metabolics (PSMs). The Nutrients constraint hypothesis argues that no one plant species can provide all the nutrient requirements of herbivores (Westoby 1978) and the Detoxification limitation hypothesis contends that mammalian herbivores are unable to reduce the detoxification loads of similar PSMs (Freeland & Janzen 1974). An alternative to overcome these limitations is to adopt a generalized feeding strategy (Wiggins et al. 2006).
Although, this animal is generalist in feeding strategy, it prefers to consume browse plant species more than the grasses. Isotopic evidence of the diets of F-hA supported that this species is a browser in Nepal (Pokharel et al. 2015). The statistically re-established hypothesis of Jarman (1974) by Brashares et al. (2000) suggests that feeding selectivity of ruminants is negatively correlated with body size and group size. Hence, smaller species require more energy per unit weight. Smaller antelopes have smaller stomachs compared to larger ruminants but have high metabolic requirements. This prohibits them from feeding large quantities of coarse grass species that are high in fiber content and low in protein content. Since highly nutritious and protein rich food is scarce, F-hA do not attain high abundances (Sharma et al. 2009). Berwick (1974) and Sharma et al. (2009) also concluded that this species was a selective feeder. The food selectivity in F-hA may result from nutritional requirements, the need to decrease fiber intake, and maximization of protein intake in order to increase digestibility.
Diet composition is the direct reflection of resource use and can provide insights into habitat utilization, habitat preferences and competitive interactions with other sympatric animals. Seasonal use of plants species shows the varieties of feeding habits. The forage selection by F-hA varied markedly during monsoon, switching their food preferences from browse plant species to grass plant species. During winter the browse to grass ratio was higher than in summer and monsoon. This shows that during summer and winter, F-hA feed largely on browse plants while during monsoon they switch their food preferences from browse plant species to grass plant species. This is because during the summer season, the grass species in the tropical forests become too coarse (Baskaran 1998) and less nutritive compared to the monsoon season (Sukumar 1989). During the winter season the cold and low moisture retards the growth of grass species and most grasses dry up becoming coarse and less nutritive. The F-hA, in turn, feed mostly on browse plant species to meet their nutritional requirements. But during monsoon, the first rain in the pre-monsoon season stimulates new grass growth, and the intercalary meristem growth of monocots is more nutritious than apical growth in browse plants (Jarman 1974). So the browse plants are substituted by grass species for food during the monsoon. This knowledge and understanding can be helpful in habitat management practices and can be implemented to enhance availability of food. The long term survival of the F-hA depends on the availability of suitable habitat hence knowing and protecting plant species utilized by the species is a significant factor in its conservation.
Accounting for the potential dietary competition of Four-horned Antelope with other sympatric ungulates in Babai valley, there is some degree of sharing of food plant species with Swamp Deer, Hog Deer, Rhinoceros and Elephant. Six of the plant species (Themeda sp., Cynodon dactylon, Imperata cylindrica, Hemarthria compressa, Mallotus philippensis and Ziziphus mauritiana) consumed by Swamp Deer, Hog Deer and Rhinoceros (Wegge et al. 2006) are also consumed by F-hA. Aegle marmelos, Bauhinia sps, Desmodium sp., Mallotus philippensis, Ziziphus mauritiana, Cynodon dactylon, Imperata cylindrica and Themeda sp. observed in the diets of Rhinoceros and Elephant (Pradhan et al. 2008) are also observed in the Four-horned Antelope diet. Interestingly, the F-hA was found to consume only one plant in common Imperata cylindrica with Barking Deer - a potential competitor. But the diet of Barking Deer was studied in Shivapuri National Park, Nepal (Nagarkoti & Thapa 2007), so generalization from the studies conducted in different landscapes and different vegetation availability may be misleading.
REFERENCES
Barcia, P., Bugalho, M.N., Campagnolo, M.L. & J.O. Cerdeira (2007). Using n-alkanes to estimate diet composition of herbivores: a novel mathematical approach. Animal 1: 141–149; http://dx.doi.org/10.1017/S1751731107340068
Baskaran, N. (1998). Ranging and resource utilization by Asian Elephant (Elephus maximus) in Nilgiri biosphere reserve, South India. PhD Thesis. Bharathi-Dasan University, Tiruchirapalli, India.
Baskaran, N., Kannan, V., Thiyagesan, K. & A.A. Desai (2011). Behavioural ecology of Four-horned Antelope (Tetracerus quadricornis Blainville, 1986) in the tropical forests of southern India. Mammalian Biology 76: 741–747; http://dx.doi.org/10.1016/j.mambio.2011.06.010
Berwick, S.H. (1974). The community of Wild ruminants in the Gir forest ecosystem, India. PhD Dissertation, University of Michigan, U.S.A.
Bhuju, U.R., P.R. Shakya, N.P. Poudel & S.S. Bajimaya (2006). Final report on the protected Area Learning Network (Phase II), EPP/FLS Project - Terai Arc Landscape, Nepal, submitted to Government of Nepal, Ministry of Forests and Soil Conservation, Department of National parks and Wildlife Conservation, and WWF Nepal Program, Kathmandu.
Blanford, W.T. (1888). The Fauna of British India including Ceylone and Burma: Mammalia. Taylor and Francis, London, U.K.
BPP (1995). Red Data Book of the fauna of the Nepal. Biodiversity Profile Project. Publication No. 4. HMG, Department of National Parks and Wildlife Conservation, Kathmandu.
Brashares, J.S., T. Garland & P. Arcese (2000). Phylogenetic analysis of co-adaptation in behavior, diet, and body size in the African Antelope. Behavioral Ecology 11(4): 452–463; http://dx.doi.org/10.1093/beheco/11.4.452
Buys, D. (1990). Food selection by eland in the western Transvaal. South African Journal of Wildlife Resources 20: 18–20.
Cavalini, P. & S. Lovari (1991). Environmental factors influencing the use of habitat in the Red Fox Vulpes vulpes. Journal of Zoology, London 223: 323–339; http://dx.doi.org/10.1111/j.1469-7998.1991.tb04768.x
Dinerstein. E. (1979). An ecological Survey of the Royal Karnali-Bardia Wildlife Reserve, Nepal. Part II: habitat/interactions. Biological conservation 16: 265-300.
DNPWC (2001). Royal Bardia National Park: Management plan 2001-2005. Kathmandu: Government of Nepal. Department of National Parks and Wildlife Conservation, Nepal.
DNPWC (2010). Fact Sheet on Banke National Park and Buffer Zone Area. Department of National Parks and Wildlife Conservation, Babarmahal, Kathmandu.
Field, C.R. (1971). Elephant ecology in the Queen Elizabeth National Park, Uganda. East African Wildlife Journal 9: 99–123; http://dx.doi.org/10.1111/j.1365-2028.1971.tb00223.x
Freeland, W.J. & D.H. Janzen (1974). Strategies in herbivory by mammals: the role of plant secondary compounds. American Naturalist 108: 269–289.
Gurung, K.K. & R. Singh (1996). Mammals of the Indian Subcontinent and Where to Watch Them. Indian Experience, Oxford, 150pp.
Harbone, J.B. (1991). The chemical basis of plant defence, pp. 45–49. In: Palo, R.T. & C.T. Robbins (eds.). Plants Defense Against Mammalian Herbiviory. CRC press, Florida.
Hofmann, R.R. (1989). Evolutionary steps of ecophysical adaptations and diversifications of ruminants. A comparative viet of their digestive system. Oecologica 78: 443–457; http://dx.doi.org/10.1007/bf00378733
Holechek, J.L. & B.D. Gross (1982). Training needed for quantifying simulated diets from fragmented range plants. Journal of Range Management 35: 644–645.
Holechek, J.L. & R. Valdez (1985). Evaluation of in vitro digestion for improving botanical estimates of mule deer faecal samples. Journal of Mammalogy 66: 574–577; http://dx.doi.org/10.2307/1380940
Holechek, J.L., M. Vavra & R.D. Pieper (1982). Botanical composition determination of range herbivore diets: a review. Journal of Range Management 35: 309–315.
IUCN. (2013). IUCN Red List of Threatened Species. Version 2013.2. <www.iucnredlist.org>. Downloaded on 18 March 2014.
Jarman, P.J. (1974). The social organization of antelope in relation to their ecology. Behaviour 48: 215–267; http://dx.doi.org/10.1163/156853974X00345
Jerdon, T. (1874). The Mammals of India. John Weldon, London.
Karki, J.B. (1998). Habitat use and population structure of Four-horned Antelope (Tetracerus quadricornis) (Blainville) in Babai Valley of Bardia National Park, Western Terai, Nepal. The Nepal Journal of Forestry 10(1): 81–90.
Klaus-Hugi, C., G. Klaus, B. Schmid & B. Konig (1999). Feeding ecology of large social antelope in the rainforest. Oecologica 119: 81–90.
Korschgen, L.J. (1971). Procedure for food-habit analysis. In: Giles, R.H. (ed.). Wildlife Techniques. The Wildlife Society, Washington, DC.
Krebs, C.J. (1999). Ecological Methodology. Addision-Wesley Longman, Menlo-park.
Krishna, Y.C., P.J. Clyne, J. Krishnaswamy & N.S. Kumar (2009). Distributional and ecological review of the Four-horned Antelope, Tetracerus quadricornis. Mammalia 73: 1–6; http://dx.doi.org/10.1515/MAMM.2009.003
Levins, R. (1968). Evolution of Changing Environments. Princeton University Press, Princeton, N.J., 120pp.
Mofareh, M.M., R.F. Beck & A.G. Schneberger (1997). Comparing techniques for determining steer diets in northern Chihuahuan Desert. Journal of Range Management 50: 27–32.
Murray, J.A. (1884). The Vertebrate Zoology of Sind. Richardson and Co., London, 424pp.
Nagarkoti, A. & T.B. Thapa (2007). Food habits of Barking Deer (Muntiacus muntjac) in the middle hills of Nepal. Hystrix It. Journal of Mammalogy (n.s.) 18(1): 77–82.
Norbury, G.L. (1988). Microscopic analysis of herbivore diets - a problem and a solution. Australian Wildlife Research 15: 51–57; http://dx.doi.org/10.1071/WR9880051
Odendaal, P.B. (1983). Feeding habits and nutrition of bushbuck in the Knysna forests during winter. South African Journal of Wildlife Resources 13: 27–31.
Owen-Smith (1993). Comparative mortality rate of male and female Kudus: the cost of sexual size dimorphism. Journal of Animal Ecology 62: 428–440; http://dx.doi.org/10.2307/5192
Parker, K.L., P.S. Barboza & M.P. Gillinggam (2009). Nutrition integrates environmental responses of ungulates. Functional Ecology 23: 57–69; http://dx.doi.org/10.1111/j.1365-2435.2009.01528.x
Pekins, P.J., K.S. Smith & W.W. Mautz (1998). The energy costs of gestation in White-tailed Deer. Canadian Journal of Zoology 76: 1091–1097; http://dx.doi.org/10.1139/z98-032
Pokharel, K.P., E. Yohannes, L. Salvarina & S. Ilse (2015). Isotopic evidence for dietary niche overlap between Barking Deer and Four-horned Antelope in Nepal. Journal of Biological Research - Thessaloniki 22: 6; http://dx.doi.org/10.1186/s40709-015-0029-0
Pokharel, K. (2010). Factors influencing the spatial distribution patterns of the Four-horned Antelope in Babai Valley, Bardia National Park, Nepal, M.Sc. Thesis. University of Freiburg, Germany.
Pokharel, K. (2012). The Four-horned Antelope: The Distribution Patterns, Resource Selection and Immediate Threats in Chitwan National Park, Nepal. Final report submitted to The Ruffords Foundation, U.K.
Pradhan, N.M.B., P. Wegge, S.R. Moe & A.K. Shrestha (2008). Feeding ecology of two endangered sympatric megaherbivores: Asian Elephant Elephas maximus and Greater One-horned Rhinoceros Rhinoceros unicornis in lowland Nepal. Wildlife Biology 14: 147–154; http://dx.doi.org/10.2981/0909-6396(2008)14[147:FEOTES]2.0.CO;2
Prater, S.H. (1971). The Book of Indian Animals. Journal of Bombay Natural. History Society Mumbai, India.
Rahmani, A.R. (2001). India, pp. 178–187. In: Mallon, D.P. & S.C. Kingswood, (comps.). Antelopes. Part 4: North Africa, The Middle East, and Asia. IUCN, Gland, Switzerland.
Rice, C.G. (1990). The status of Four-horned Antelope Tetracerus quadricornis. Journal of Bombay Natural History Society 88: 63–66.
Roberts, T.J. (1997). The Mammals of Pakistan. Ernest Benn Limited, London, 361pp.
Rodgers, W.A. & H.S. Panwar (1988). Planning Wildlife Protected area Network in India. FAO Field Document Dehradun 1(9): 341.
Sandoval, L., J. Holechek, J. Biggs, R. Valdez & D. Vanleeuwen (2005). Elk and Mule Deer diets in north-central New Mexico. Rangeland Ecology and Management 58: 366–372.
Sankala, K. (1977). Tiger! The Story of the Indian Tigers. Rupa and Co., Collins, London, 220pp.
Sharma, K. (2006). Distribution, status, ecology and behaviour of the Four-horned Antelope Tetracerus quadricornis, pp. 72–82. In: Sharma, K., A.R. Rahmani & R.S. Chundawat (2009) (eds.). Natural History Observations of the Four-horned Antelope Tetracerus quadricornis. Journal of the Bombay Natural History Society 106(1): 72–82.
Sharma, K. & A.R. Rahmani (2005). Four Horned Antelope or Chowsingha (Tetracerous quadricornis, Blainville, 1816). ENVIS bulletin (Wildlife and Protected Areas): Ungulates of India.
Sharma, K., R.S. Chundawat, J.V. Gruisen & A.R. Rahmani (2013). Understanding the patchy distribution of Four-horned Antelope Tetracerus quadricornisin a tropical dry deciduous forest in Central India. Journal of Tropical Ecology, Available on CJO 2013; http://dx.doi.org/10.1017/S0266467413000722
Sharma, K., A.R. Rahmani & R.S. Chundawat (2005). Ecology and Distribution of Four-horned Antelope Tetracerus quadricornis in India. Final report-DST, Bombay Natural History Society, Mumbai, India, 70pp.
Sharma, K., A.R. Rahmani & R.S. Chundawat (2009). Natural History Observations of the Four-horned Antelope Tetracerus quadricornis. Journal of the Bombay Natural History Society 106(1): 72–82.
Shrestha, K.K., P.K. Jha & J.K. Ghimire (1997). Final technical report: Plant diversity Analysis and Evaluation of conservation measures in the Royal Bardia National Park, Nepal. WWF Nepal Program, Kathmandu, Nepal.
Shrestha, T.K. (2001). Nepal, pp. 188–191. In: Mallon, D.P. & S.C. Kingswood (eds.). Antelopes. Part 4: North Africa, the Middle East, and Asia. Global Survey and Regional Action Plans, IUCN, Gland.
Shipley, L.A. (1999). Grazers and browsers: how digestive morphology affects diet selection, pp. 20–27. In: Launchbaugh, K.L., K.D. Sanders & J.C. Mosley (eds.). Grazing Behaviour of Livestock and Wildlife. Idaho Forest, Wildlife and Range Expeditions Station Bulletin, University of Idaho, Moscow.
Solanki, G.S. & R.M. Naik (1998). Grazing interactions between wild and domestic herbivores. Small Ruminant Research 27: 231–235.
Steinheim, G., P. Wegge, J.I. Fjellstad, S.R. Jnawali & R.B. Weljadi (2005). Dry season diets and habitat use of sympatric Asian Elephants (Elephus maximus) and Greater One-horned Rhinoceros (Rhinocerus unicornis) in Nepal. Journal of Zoology 265: 377–385.
Sukumar, R. (1989). The Asian Elephant: Ecology and Management. Cambridge University Press, Cambridge, UK.
Vavra, M. & J.L. Holechek (1980). Factors influencing microhistological analysis of herbivore diets. Journal of Range Management 33: 371–374.
Wegge, P., A.K. Shrestha & S.R. Moe (2006). Dry season diets of sympatric ungulate in lowland Nepal: competition and facilitation in alluvial tall grasslands. Ecological Research 21: 698–706.
Westoby, M. (1978). What are the biological bases of varied diets? American Naturalist 112: 627–631.
Wiggins, N.L., C. McArthur & N.W. Davies (2006). Diet switching in a generalist mammalian folivore: fundamental to maximizing intake. Oecologia 147: 650–657; http://dx.doi.org/10.1007/s00442-005-0305-z