Habitat use and relative abundance of the Spotted Paca Cuniculus paca(Linnaeus, 1766) (Rodentia: Cuniculidae) and the Red-rumped Agouti Dasyprocta
leporina (Linnaeus, 1758) (Rodentia: Dasyproctidae) in Guatopo National
Park, Venezuela
Elinor Jax 1, Sofía Marín 2,
Adriana Rodríguez-Ferraro 3 & Emiliana Isasi-Catalá 4
1 Institute
of Biology, Lund University, SE-221 00 Lund, Sweden
2,4 Laboratorio
de Conservación y Manejo de Fauna Silvestre, Universidad Simón Bolívar, Valle
de Sartenejas, Código postal 1020. Baruta, Caracas, Venezuela
3 Departamento
de Estudios Ambientales, Universidad Simón Bolívar, Valle de Sartenejas, Código
postal 1020, Baruta, Caracas, Venezuela
1 elinorjax@gmail.com (corresponding author), 2sofimarinwikander@gmail.com, 3 rodrigueza@usb.ve,4 zmiliana@gmail.com
Abstract: The Spotted Paca Cuniculus paca and the
Red-rumped Agouti Dasyprocta leporina are affected by habitat loss and
hunting. In Venezuela, their
conservation status is unknown, even within protected areas. The objective of this study was to estimate
the relative abundance, activity patterns, habitat use, and effect of human
activities on these species in Venezuela. To achieve this, 26 camera-trap stations (20.8km2) were
established in Guatopo National Park between February and April 2011,
characterization of the habitat was undertaken and occupancy models were
created. The relative abundance of the
Spotted Paca was 1.62 captures/100trap-nights, with a fully nocturnal activity
pattern. The relative abundance of the
Red-rumped Agouti was 2.32 captures/100trap-nights, with a pronounced diurnal
activity pattern. The occupation probability of the Red-rumped Agouti (0.61 SE
0.02) was higher than that of the Spotted Paca (0.27 SE 0.02). Spotted Pacas
were mainly found in areas with mature forest and high tree density, whereas
the Red-rumped Agoutis were most
frequently found in valleys with little disturbed forest. A positive correlation was found between
illegal hunting activities and areas occupied by the Spotted Paca. It is important to strengthen the park
control measurements to reduce illegal hunting of Spotted Pacas.
Keywords: Camera traps, Cuniculus paca, Dasyprocta
leporina, habitat use, human activities, illegal hunting, occupancy models,
protected area.
Abbreviations: AICc - Adjusted Akaike Information Criteria; LV -
Amount of leaves in decomposition; Bψ - Beta coefficient for occupancy
variables; Bp - Beta coefficient for detection variables; p - Detection
probability; hi - Detection/non-detection histories; DP - Detection of
predators; DLP - Detection of the two largest predators; WA - Distance to the
closest water source; DR - Distance to the main road; HA - Frequency of human
activity; FT - Forest type; GNP - Guatopo National Park; GV - Ground vegetation
coverage; HV - Horizontal vegetation coverage; IUCN - International Union for
Conservation of Nature; LC - Least Concern; c - Model fit; wi - Model weight; δ
- Number of parameters; t-n - Number of trap-nights; C - Percentage of activity
crepuscular; D - Percentage of activity
diurnal; N - Percentage of activity nocturnal; Ψ - Probability that a site is
occupied; RAbundance - Relative abundance; RaP - Relative abundances of
predators; RaLP - Relative
abundances of the two largest predators; sign - Significance of the model;
RaAgouti - Site specific relative abundance of the Red-rumped Agoutis; RaPaca - Site specific
relative abundance of the Spotted Paca; TO – Topography, TD - Tree density; VV
- Vertical vegetation coverage.
doi: http://dx.doi.org/10.11609/JoTT.o3915.6739-49 | ZooBank: urn:lsid:zoobank.org:pub:47CAD9C8-EF5E-4FDE-A755-FC9914C44208
Editor: Giovanni Amori, CNR - Institute of Ecosystem Studies, Roma, Italy. Date of publication: 26 January 2015
(online & print)
Manuscript details: Ms # o3915 |
Received 15 January 2014 | Final received 29 November 2014 | Finally accepted
21 December 2014
Citation: Jax, E., S. Marín, A. Rodríguez-Ferraro & E. Isasi-Catalá (2015). Habitat
use and relative abundance of the Spotted Paca Cuniculus paca (Linnaeus,
1766) (Rodentia: Cuniculidae) and the Red-rumped Agouti Dasyprocta leporina(Linnaeus,1758) (Rodentia: Dasyproctidae) in Guatopo National Park,
Venezuela. Journal of Threatened Taxa 7(1):6739–6749; http://dx.doi.org/10.11609/JoTT.o3915.6739-49
Copyright: © Jax et al. 2015.
Creative Commons Attribution 4.0 International License. JoTT allows
unrestricted use of this article in any medium, reproduction and distribution
by providing adequate credit to the authors and the source of publication.
Funding: This study was supported by Fundación para la
Defensa de la Naturaleza (FUDENA) and IdeaWild.
Competing Interest: The authors declare
no competing interests.
Author
Details: Elinor Jax has a
BSc in Biology and a MSc in Ecology from Lund University. She has been involved
in projects related to ecology, conservation, population genetics, and the
effect of infectious diseases in wildlife populations. Emiliana
Isasi-Catalá is a Doctor in Biological Sciences at Universidad Simón
Bolívar. Her research area is the ecology and conservation of terrestrial
vertebrates. She is particularly interested in the ecological study of
surrogate species and species presenting conflicts with humans, such as big
cats and their prey. Sofía Marín is a biologist at
Universidad Simón Bolívar in Venezuela. Her research focuses on the ecology and
population biology of Lowland Tapirs Tapirus terrestris in Cordillera de la
Costa. She is interested in the conservation of wildlife and socio-ecological
interactions that allow sustainable management of natural resources. Adriana Rodríguez-Ferraro got her PhD at
the University of Missouri-St- Louis and is currently an Associate Professor at
Universidad Simón Bolívar. Her research area is on ecology and conservation of
threatened and restricted-range terrestrial vertebrates, especially birds.
Author Contribution: EIC and
SMW created the sampling design. EJ, EIC and SMW collected and organized the
data. EJ and EIC analyzed the data. EJ, EIC and ARF prepared and reviewed the
article.
Acknowledgements: We are
grateful to the people working in Guatopo National Park who made this project
possible. Particularly, we would like to
thank Habi Veroes for his invaluable knowledge about Guatopo, and for sharing
this knowledge with us. We would also like to give a special thanks to all
field assistants who followed us to Guatopo when we needed them the most.

INTRODUCTION
The Spotted Paca Cuniculus paca(Linnaeus, 1766) and the Red-rumped Agouti Dasyprocta leporina(Linnaeus, 1758) are large terrestrial rodents that inhabit a variety of
habitats from northern Argentina to southern Mexico (Emmons & Feer 1990;
Nowak 1991; Perez 1992). Their diet
includes fruits, seeds and nuts (Eisenberg 1989), making them important seed
dispersers in the tropics (Forget 1994; Henry 1999). Both species are classified as Least Concern
(LC) by the IUCN Red List, mainly because of their wide distribution and
occurrence within protected areas (Emmons & Reid 2008; Queirolo et al.
2008).
Both species have a wide
distribution in Venezuela and are considered common, however in recent decades
they have been negatively affected by habitat loss as well as subsistence and
commercial hunting (Silva & Strahl 1994, 1996; Linares 1998). The Spotted Paca is subjected to a high
hunting pressure in Venezuela (Images 1–2) because of its high value meat
(Silva & Strahl 1994). Its numbers may have declined or it might even have
been locally extirpated at some locations where it was previously abundant, as
has been reported in other parts of its distribution (Emmons & Feer 1990;
Nowak 1991). Nevertheless, neither the
Spotted Paca nor the Red-rumped Agouti
are considered threatened by the Venezuelan government, and thus they are not
protected from hunting (Venezuela 1996a,b) or included in the Venezuelan Red
List (Rodríguez & Rojas-Suárez 2008). The conservation status of these rodents is of great concern since no
recent studies have been conducted in Venezuela, hence information on their
abundance, habitat use, and threats is limited.
With the purpose of improving this
knowledge we conducted a study in Guatopo National Park (GNP) using camera
traps and occupancy models. Camera-traps
greatly facilitate the study of cryptic mammals in remote areas (Karanth &
Kumar 2002; Srbek-Araujo & Garcia Chiarello 2005; Kays & Slauson 2008;
Tobler et al. 2008; Rovero et al. 2013) and have, therefore, become one of the
most common tools for studying mammals in the Neotropics (Kuroiwa
& Ascorra 2002; Polisar et al. 2003; Maffei et al. 2005; Noss et al. 2006;
Ríos-Uzeda et al. 2007; Schipper 2007; Blake et al. 2012). Some
camera trap studies have reported data on the relative abundance (Maffei et al.
2002; de Souza Martins et al. 2007; Tobler et al. 2008), activity patterns
(Blake et al. 2012), and occupation probability (Ahumada et al. 2013;
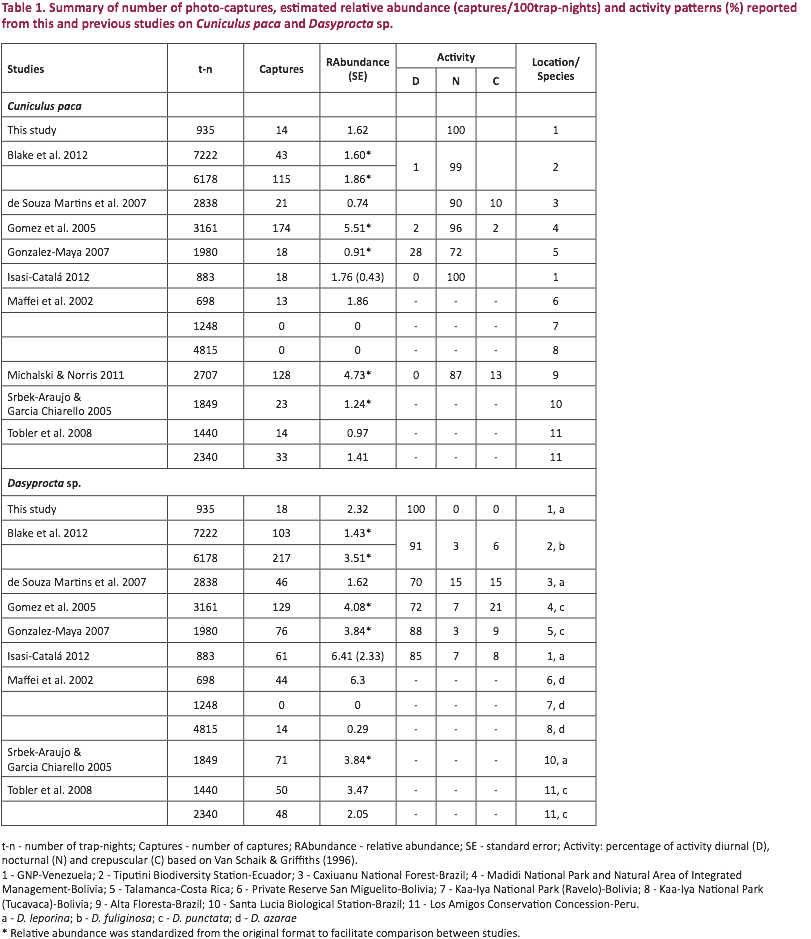
Isasi-Catalá 2013) of the Spotted Paca and the Red-rumped Agouti (Table 1).
Occupancy models are primarily
used when studying species that are difficult to detect (MacKenzie et al. 2002;
Linkie et al. 2007; Ahumada et al. 2013). From these models one can estimate the detection probability (p) and the
probability that a site is occupied (Ψ) by a particular species based on the
analysis of their detection/non-detection histories (hi) (MacKenzie et al.
2002; Royle & Nichols 2003; MacKenzie et al. 2006). By combining this data with environmental
variables from the study area, habitat use can be evaluated (MacKenzie et al.
2002; Royle & Nichols 2003; MacKenzie et al. 2006).
The objective of this study was to
estimate the relative abundance, activity patterns, habitat use, and effect of
human activities on the Spotted Paca and Red-rumped Agouti in a protected area of the Cordillera de la Costa to
assess the conservation status of these species in Venezuela. To do so we conducted a camera trap survey in
Guatopo National Park (GNP), where previous studies have reported several cases
of illegal hunting of the Spotted Paca (Silva & Strahl 1996; Isasi-Catalá
2012).
MATERIAL AND METHODS
Study area and data collection
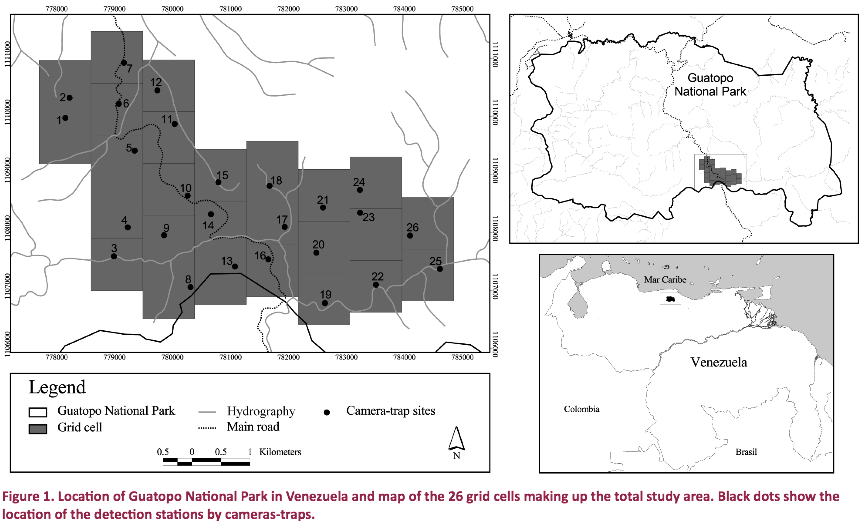
GNP (1,224.64km2, Fig.
1), is located in the Cordillera de la Costa in northern Venezuela (10.03N
& 66.41W) (Venezuela 1958). The park
is intersected by numerous rivers and mountain ranges with steep inclines and
elevations ranging from 200 to 1,430 m (Yerena 1985; MARNR 1992). The climate
is warm and humid with an annual rainfall between 1,400–2,800 mm and an average
annual temperature between 18–32°C (Castillo & Aponte 2004). Predominant vegetation types within the park
are rain forests, cloud forests, and semi deciduous forests (Yerena 1985;
Perera-Romero 2006). The national park
lies within the most populated regions in the country and is considered
vulnerable due to human activities (Castillo & Aponte 2004; Blanco &
Yerena 2006).
The study was conducted in the
south-western part of GNP (Fig. 1) between February and April 2011. The study area (20.8km2) was
divided into 26 adjacent grid cells (Fig. 1), each containing a photo-capture
station: a single camera-trap (Bushnell Trophy Cam, Moultrie Game Spy i65, or
Moultrie Game Spy I-40) and an attractor (perfume Chanel N°5, Perfume
Factory). Photo-capture stations were
established on tracks or other sites where signs of wildlife activity were
observed. Two types of data were
obtained from camera-traps: count of capture events and detection/no detection
of individuals (Karanth & Kumar 2002; MacKenzie et al. 2006; Kays &
Slauson 2008). As the Spotted
Paca and the Red-rumped Agouti cannot be
individually differentiated multiple photos taken within a short period of time
might capture the same individual, and so could lead to an overestimation of
the abundance of the species. For the count of capture events, photos taken
within a 24h period were therefore considered the same event if the photo or
video did not clearly show that it was a different individual: male/female,
adult/juvenile. This conservative
approach follows the methodology used in previous studies (Maffei et al. 2002;
González-Maya 2007; Isasi-Catalá 2012).
To assess the rodents habitat use,
a characterization of the habitat was conducted in each grid cell according to
their area of action (3rd order selection) (Johnson 1980; Dungan et al. 2002;
Boyce et al. 2003). At each station, eight
environmental variables were determined: ground (GV), horizontal (HV), and
vertical vegetation (VV) coverage, amount of leaves in decomposition (LV), tree
density (TD), distance to the closest water source (WA), topography (TO), and
forest type (FT). GV, HV, and VV was
estimated with the program CobCal V.1.0© (Ferrari et al. 2009) based on
photographs. LV was measured in cm and TD was calculated according to the
Centre Point Square Method (Mostacedo & Fredericksen 2000). WA was categorized as either present within
a distance less than 50m, between 50–200 m or greater than 200m from the
station, based on the target species estimated dispersal distance (Beck-King et
al. 1999). TO was categorized as
valley, peak or intermediate and FT was classified as mature or little
disturbed forest, based on the classification by Yerena (1985).
Other factors, such as predation
(Abramsky et al. 1996; Heithaus & Dill 2002; Sundell et al. 2004),
competition (Wasserberg et al. 2006; Head et al. 2012), and human activities
(Franklin et al. 2002) might also influence species habitat use. Based on records obtained from camera-traps
six additional variables were therefore determined: the relative abundances
(RaP) and detection (DP) of predators (P: Puma concolor, P. onca, Leopardus
pardalis, L. wiedii, and L. tigrinus), the relative
abundances (RaLP) and detection (DLP) of the two largest predators: (LP: P.
concolor, P. onca), and the site specific relative abundance of each target
species (RaAgouti and RaPaca). Further,
to evaluate the impact of human activities evidence of illegal hunting and
logging recorded in this and previous studies in GNP (Isasi-Catalá 2009, 2012)
were used to calculate the frequency of human activity (HA) for each grid cell.
Additionally, distance to the main road (DR) was extracted from maps in ArcView
3.2® (ESRI Inc., Redlands, CA).
Data analysis
Mean relative abundance and site
specific relative abundances were calculated, as the number of photo-captures
made per 100 trap-nights, for each species. Site specific relative abundances were also calculated and compared
between the species using a Mann-Whitney U test in SPSS20.0© (SPSS Inc.,
Chicago, IL). The activity pattern for
each species was divided per hour and compared with the Morisita-Horn similarity
index (Horn 1966) using EstimateS (Colwell 2001). The uniformity of its distribution was then
evaluated with the Kolmogorov-Smirnov test in SPSS20.0© (SPSS Inc., Chicago,
IL). The activity pattern was classified as diurnal (06:00–17:00), nocturnal
(18:00–05:00) or crepuscular (05:00–06:00 and 17:00–18:00) (Van Schaik &
Griffiths 1996) for comparison with other studies.
Species habitat use was assessed
through occupancy models (MacKenzie et al. 2006), using the program Presence
2.4 (Hines 2006). Detection histories
for each species were built from records obtained by camera traps using a
binary system with zeros indicating lack of detections and ones indicating
detections (MacKenzie et al. 2002; MacKenzie et al. 2006). Ψ was estimated from the variables GV, HV,
VV, LV, TD, WA, TO, FT, Ra_paca, Ra_agouti, RaP, RaLP, HA and DR, and p from
RaP, DP, RaLP, DLP, Ra_paca, Ra_agouti. Models were selected based on model goodness of fit, adjusted Akaike
Information Criteria (AICc), AIC model weight, and the dispersion of the models
(MacKenzie et al. 2006; Linkie 2008). The top ranked occupancy models (ΔAICc < 3, MacKenzie et al.
2006) were used to determine Ψ of each species for each grid. Ψ were compared
between the species with a Mann-Whitney U test implemented in SPSS20.0© (SPSS
Inc., Chicago, IL). The variables
included in these models were used to determine the characteristics of the
habitat used by each species, based on the beta coefficient (MacKenzie et al. 2006):
Bψ for occupancy variables and Bp for detection variables.
RESULTS
Relative Abundance and Activity Patterns
A total of 935 trap-nights were
performed, during which there were 14 independent capture events of Spotted
Pacas at 6 sites and 18 capture events of Red-rumped Agoutis at 12 sites (Table 1, Video 1). Mean relative abundance was calculated for
each species, resulting in 1.62 captures/100trap-nights for the Spotted Paca
and 2.32 captures/100trap-nights for the Red-rumped Agouti (Table 1). No
significant difference was found between the species site specific relative
abundances (Mann-Whitney U = 274.50; Z = -1.37; P = 0.17, n = 52).
All captures of the Spotted Paca
were made between 21:00 and 05:00 (nocturnal), whereas all except one capture
of the Red-rumped Agouti were made between
06:00 and 17:00 (diurnal, Table 1, Fig. 2). The number of captures was not uniformly distributed over the day for
either species (Kolmogorov-Smirnov; Spotted Paca: Z = 3.27; P < 0.01; n =
24; Red-rumped Agouti: Z = 3.06; P <
0.01; n = 24), and no overlap between the activity hours of the two species was
observed (Morisita-Horn similarity index = 0%).
Habitat use and impact of human activities
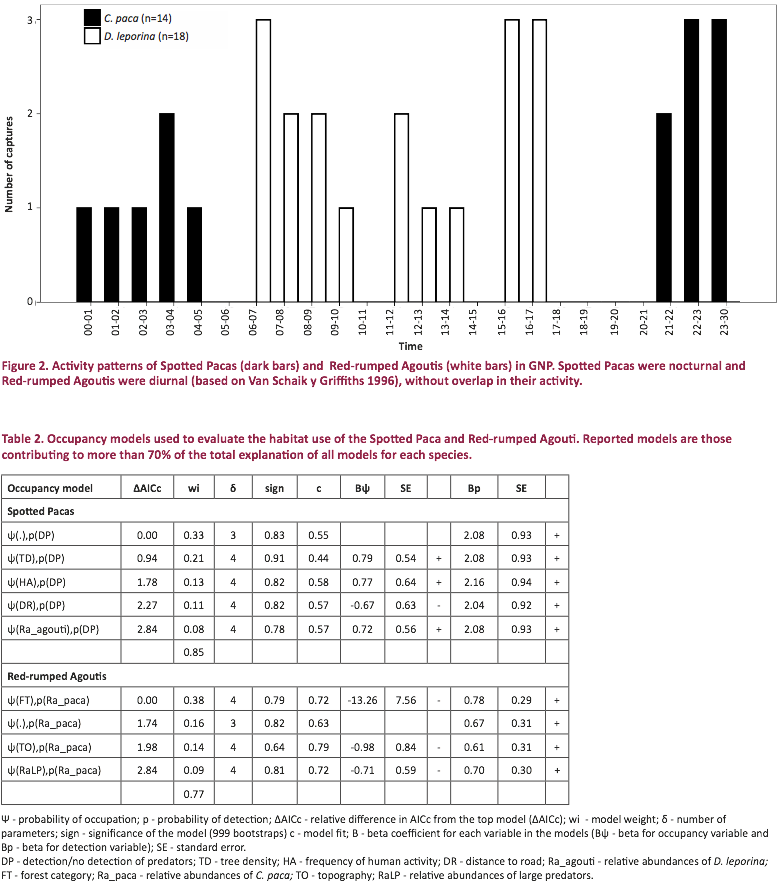
According to occupancy models
selected (covering >70% of the total explanation power, Table 2), mean
occupation probabilities for the Spotted Paca and the Red-rumped Agouti were 0.27 (SE 0.02) and
0.61 (SE 0.02), respectively. The site
specific occupation probability was significantly higher for the Red-rumped Agouti than for the Spotted Paca
(Mann-Whitney U = 16.00; Z = -5.91; P < 0.01; n = 52). In GNP, the Spotted Paca was associated with
areas with high density of trees (BψTD = 0.79, SE = 0.54 Table 2),
located at greater distances from the main road (BψDR = -0.67, SE =
0.63), and characterized by a high presence of predators (BpDP >
2.04, SE < 0.94), as well as high abundance of the Red-rumped Agouti (BψRa_agouti =
0.72, SE = 0.56). Areas with a high probability of being occupied by the
Spotted Paca also had high frequency of illegal human activities (BpHA= 0.77, SE = 0.64 Table 2). Red-rumped
Agoutis were strongly associated with little disturbed forest (BψFT= -13.26, SE = 7.56, Table 2) and valleys (BψTO = -0.98, SE =
0.84). Sites occupied by the Red-rumped Agouti had a high presence of Spotted
Pacas (BpRa_paca > 0.61, SE < 0.31) but low abundance of large
predators (BψRaLP = -0.71, SE = 0.59, Table 2). No correlation was found between species
occupancy and the other variables studied (like vegetation cover or distance to
watercourses).
DISCUSSION AND CONCLUSION
Relative Abundance and Activity Pattern
Relative abundance is a measure
based on sampling effort and differences in methodology therefore makes it
problematic to compare results between studies (Srbek-Araujo & Garcia
Chiarello 2005; Tobler et al. 2008; Michalski & Norris 2011). Still, it is important to highlight that the
relative abundances estimated for both species in our study are within the
range reported from previous camera trap studies (Table 1). In a study in Caxiuanu National Forest,
Brazil, lower relative abundances were reported for both species (de Souza
Martins et al. 2007). In contrast, a
higher relative abundance was estimated in Madidi National Park, Bolivia, for
the Spotted Paca (Gomez et al. 2005), and in Santa Lucia Biological Station in
Brazil (Srbek-Araujo & Garcia Chiarello 2005), as well as in a previous
study in GNP (Isasi-Catalá 2012) for the Red-rumped Agouti. In the last two studies the Red-rumped Agouti was one of the mammals with
highest relative abundance. Similar
relative abundances have been estimated for other species from the genus Dasyprocta,
such as D. puntata in Peru (Tobler et al. 2008) and D. azarae in
Bolivia (Maffei et al. 2002) (Table 1).
Another variable that has been
suggested as appropriate surrogate for evaluating abundance of a species is the
occupation probability (MacKenzie & Nichols 2004). In our study, the Red-rumped Agouti had a higher occupation
probability than the Spotted Paca. These
large rodents have similar characteristics and requirements (Eisenberg 1989;
Nowak 1991), thus one could expect to find them in similar abundances. It is possible that hunting pressure on the
Spotted Paca has affected its occupation probability. In the 1990s the Spotted Paca was listed as
one of the species suffering from the highest hunting pressure in GNP, though
during this period these activities were primarily taking place close to the
park limit (Silva & Strahl 1994, 1996). Today there is also a significant
hunting pressure on the species in the central part of the park, mainly close
to the large rivers located in this area (Isasi-Catalá 2012).
The activity patterns obtained in
this study coincide with those reported in other studies (Table 1). In Tiputini Biodiversity Station (Blake et
al. 2012), Talamanca (González-Maya 2007), Madidi National Park (Gomez et al.
2005) and GNP (Isasi-Catalá 2012) results indicate that the Spotted Paca is
nocturnal (Van Schaik & Griffiths 1996), whereas agoutis showed diurnal
activity patterns. Ecological factors,
such as predation (Sundell et al. 2004), resource availability (Tarnaud 2006),
and competition (Wasserberg et al. 2006) can influence the activity pattern of
a species. According to a study in Bolivia
direct competition between the Spotted Paca and the Central American Agouti (D.
punctata) is avoided through temporal separation (Gomez et al. 2005). Our results suggest that this is true for the
Spotted Paca and the Red-rumped Agouti because we did not find an overlap
between the species active hours.
Camera traps have proved efficient
tools for studying cryptic medium sized mammals like the Spotted Paca and
the Red-rumped Agouti (Srbek-Araujo
& Garcia Chiarello 2005). However,
to accurately assess the conservation status of these species it is crucial
that this sampling method is implemented with designs that give comparable and
unbiased population estimators. One of
the main limitations of using camera-traps for the study of these rodents is
the uncertainty in the identification of independent events. In studies where animals can be
differentiated due to individual markings each capture can be included in
abundance estimations (Karanth & Kumar 2002; Maffei et al. 2002; Noss et
al. 2006; Kays & Slauson 2008; Isasi-Catalá 2012; Negroes et al.
2012). It was, however, not possible
to differentiate between individuals neither for the Spotted Paca nor the
Red-rumped Agouti (Video
1). Further, both the Spotted Paca and
Red-rumped Agouti are territorial species (Linares 1998) and it is therefore
likely that captures from the same camera within a short period of time are of
the same individual inhabiting that particular territory. We therefore chose to take a more
conservative measurement and allow for the possibility that two captures within
a 24 hour period might be of the same individual on its daily run. This has been done in earlier studies of
species without distinctive markings as well (Maffei et al. 2002; González-Maya
2007; Isasi-Catalá 2012). Some studies
have used shorter intervals (even 30 min) to separate events captured with
camera-traps (Blake et al. 2012). In
total only one capture was eliminated in our study due to this conservative
approach.

Habitat use and impact of human activities
In accordance with previous
studies (Goulart et al. 2009; Ahumada et al. 2013), the Spotted Paca was mainly
found in mature forest with high tree density. The Red-rumped Agouti, on the
other hand, was mainly encountered in little disturbed forested areas, which
are often contiguous with the mature forest in the park. At present, more than 85% of the park is
covered by these types of habitats, with a high variety and abundance of plants
that provide food for many species (Isasi-Catalá 2012). High presence of palms in the park could also
be a key factor in determining habitat use of the Spotted Paca and Red-rumped Agouti, since it has been reported
that their fruits and seeds are an important food source for these species
(Emmons & Feer 1990; Ahumada et al. 2013).

Availability of watercourses is
probably an important factor for these species, particularly for the Spotted
Paca (Emmons & Feer 1990; Perez 1992; Goulart et al. 2009). However, it is possible that the pattern is
not evident at GNP as the park is full of rivers and water ponds (Yerena
1985). Another variable that is often
related to the presence of water bodies is topography (Perera-Romero 2006), we
therefore expected to find the target species in valleys rather than at peaks,
as was the case for the Red-rumped
Agouti. Similar results have been
reported from another study, indicating that the Red-rumped Agouti has a preference for
occupying lowland forest (Ahumada et al. 2013).
Although the Spotted Paca and the
Red-rumped Agouti were often separated by space and time, some of the sites
with high abundance of one species in GNP were also inhabited by the other
species. In La Selva in Costa Rica it
has been reported that although the two target species of this study may
compete for resources, the Spotted Paca can specialize in fruits whereas
the Red-rumped Agouti mainly forages on
seeds, thereby decreasing competition for food resources (Ahumada et al. 2013). Both species might however compete with other
species for these same resources, such as the Collared Peccary Pecari tajacu(Ahumada et al. 2013), which is abundant in GNP (Isasi-Catalá 2012).
The relationship between predators
and prey are not always straightforward and might therefore be difficult to
interpret. In GNP, sites that were occupied by Spotted Pacas had a high
presence of the five predators. In contrast, sites that were occupied by
the Red-rumped Agouti were negatively
correlated with the abundance of the large predators in the park: jaguar and
puma. This might be a result of the
predators’ choice of prey as well as their activity pattern in the area. Several studies imply that the Spotted Paca
is one of the most common prey for large predators, especially jaguars
(Aliaga-Rossel et al. 2006; Weckel et al. 2006). A previous study on habitat use of Jaguars in
GNP did indicate a high predation pressure by the jaguar also on the Red-rumped Agouti (Isasi-Catalá 2012). This pattern was not observed in the case of
Spotted Pacas, and could explain the relationship with predators obtained in
our study.
According to the occupancy models,
areas occupied by Spotted Pacas had a higher frequency of human activities than
areas without any signs of Spotted Pacas. This could indicate that hunters select areas with a high presence of
Spotted Pacas in GNP as the species is one of the most sought after due to its
commercial value (Silva & Strahl 1994, 1996). Recent signs of a Spotted Paca kill were found
during the study in an area where no other observation of Spotted Pacas was
made (Images 1–2). The models did not
indicate a relationship between human activity and occupation of the Red-rumped Agouti suggesting that this
species might be a secondary prey for the poachers in the park. The effect of roads on the occupation of
Spotted Pacas in GNP is similar to that reported in the Brazilian Atlantic
Forest, where Spotted Pacas were recorded more often on narrow trails than on
wider trafficked roads (Goulart et al. 2009). The main road that intersects GNP is associated with major hunting
activities within the park (Isasi-Catalá 2012), being the only way for hunters
to enter, exit, and transport their prey out of the park.
The occupancy models developed in
this study proved to be useful tools for assessing habitat use and occupation
probability of the Spotted Paca and the Red-rumped Agouti in GNP. We
believe that these models have great potential for studying temporal and
spatial patterns of many species, especially those with low detectability, even
at large scales. The use of camera trap
data in this type of models could be an efficient method for generating
reliable results at low cost in time and effort (Srbek-Araujo & Garcia
Chiarello 2005; MacKenzie et al. 2006; Tobler et al. 2008; Ahumada et al.
2013).
Implications for species conservation
As habitat loss and fragmentation
have been recognized as threats for the target species of this study (Emmons
& Feer 1990; Nowak 1991), the protection of mature and dense forests must
be one of the main strategies to ensure the long-term survival of the
species. The creation of GNP has indeed
resulted in protection and recovery of important forest cover in Cordillera de
la Costa (Isasi-Catalá 2012), one of the most threatened habitats in Venezuela
(Yerena 1985; Castillo & Aponte 2004). However, it is essential to strengthen the control of human activities
that are currently generating loss of forest cover by the park limits
(Isasi-Catalá 2012) to ensure the conservation of these species and their habitat. Another great concern for the survival of the
target species in GNP is the impact of illegal hunting. A more strict control of who enters the park
might be required to reduce the hunting pressure on the Spotted Paca and other
game animals within the park.
We believe that it is crucial to
increase the information on the status of the Spotted Paca and Red-rumped
Agouti in GNP and others areas of Venezuela in order to generate more
appropriate conservation guidelines for them. Information gathered in this and
future studies should be used to increase the awareness of the general public
and policy makers about the current situation and threats to wildlife in the
country.
REFERENCES
Abramsky, Z., E. Strauss, A. Subach, A. Riechman & B.P. Kotler
(1996). The effect of Barn Owls (Tyto
alba) on the activity and microhabitat selection of Gerbillus allenbyiand G. pyramidum. Oecologia 105(3): 313–319; http://dx.doi.org/10.1007/BF00328733
Ahumada, J.A., J. Hurtado & D. Lizcano (2013). Monitoring the status and trends of tropical forest
terrestrial vertebrate communities from camera trap data: a tool for
conservation. PLoS ONE 8(9): e73707; http://dx.doi.org/10.1371/journal.pone.0073707
Aliaga-Rossel, E., R.S. Moreno, W.K. Roland & J. Giacalone (2006). Ocelot (Leopardus pardalis) predation on
Agouti (Dasyprocta punctata). Biotropica 38(5): 691–694; http://dx.doi.org/10.1111/j.1744-7429.2006.00192.x
Beck-King, H., O. von Helversen & R. Beck-King (1999). Home range, population density, and food resources
of Agouti paca (Rodentia: Agoutidae) in Costa Rica: A study using
alternative methods. Biotropica 31(4): 675–685; http://dx.doi.org/10.1111/j.1744-7429.1999.tb00417.x
Blake, J.G., D. Mosquera, B.A. Loiselle, K. Swing, J. Guerra & D.
Romo (2012). Temporal activity patterns of
terrestrial mammals in lowland rainforest of eastern Ecuador. Ecotropica18: 137–146.
Blanco, C. & E. Yerena (2006). Metodología de Evaluación del Estado de Riesgo de un Sistema de Áreas
Protegidas. 1er Taller Nacional para el Fortalecimiento de la Efectividad de
Manejo en Áreas Protegidas de Venezuela, Caracas, Venezuela.
Boyce, M.S., J.S. Mao, E.H. Merrill, D. Fortin, M.G. Turner, J. Fryxell
& P. Turchin (2003). Scale and
heterogeneity in habitat selection by Elk in Yellowstone National Park. Ecoscience10(4): 421–431.
Castillo, R. & C. Aponte (2004). Perfil de Área Protegida - Venezuela Parque Nacional Guatopo.
ParksWatch, Caracas, Venezuela.
Colwell, R. (2001). EstimateS:
statistical estimation of species richness and shared species from samples.
Version 8.0.0. University of Connecticut, Storrs, US.
de Souza Martins, S., J.G. Sanderson & J. de Sousa e Silva-Junior
(2007). Monitoring mammals in the
Caxiuanu National Forest, Brazil - First results from the Tropical Ecology,
Assessment and Monitoring (TEAM) program. Biodiversity and Conservation16(4): 857–870.
Dungan, J.L., J.N. Perry, M.R.T. Dale, P. Legendre, S. Citron-Pousty,
M.J. Fortin, A. Jakomulska, M. Miriti & M.S. Rosenberg (2002). A balanced view of scale in spatial statistical
analysis. Ecography 25(5): 626–640; http://dx.doi.org/10.1034/j.1600-0587.2002.250510.x
Eisenberg, J.F. (1989). Mammals
of the Neotropics: The northern Neotropics, Volume 1: Panama, Colombia,
Venezuela, Guyana, Suriname, French Guiana. University of Chicago Press,
Chicago, US, 550pp.
Emmons, L. & F. Reid (2008). Dasyprocta leporina. IUCN Red List of Threatened Species
Version 2012.2 Available online 26 March 2013; www.iucnredlist.org
Emmons, L.H. & F. Feer (1990). Neotropical Rainforest Mammals: A Field Guide. University of
Chicago Press, Chicago, US, 281pp.
Ferrari, D.M., O.R. Pozzolo & H.J. Ferrari (2009). Software de Estimación de Cobertura Vegetal -
CobCal. Version 1.0. Estación Experimental Agropecuaria INTA Concepción del
Uruguay, Entre Ríos, Argentina.
Forget, P.M. (1994). Recruitment
pattern of Vouacapoua americana (Caesalpiniaceae), a rodent- dispersed
tree species in French Guiana. Biotropica 26(4): 408–419.
Franklin, A.B., B.R. Noon & T.L. George (2002). What Is Habitat Fragmentation? Studies in Avian
Biology 25: 20–29.
Gomez, H., R.B. Wallace, G. Ayala & R. Tejada (2005). Dry season activity periods of some Amazonian
mammals. Studies on Neotropical Fauna and Environment 40(2): 91–95; http://dx.doi.org/10.1080/01650520500129638
González-Maya, J.F. (2007).
Densidad, uso de hábitat y presas del Jaguar (Panthera onca) y el
conflicto con humanos en la región de Talamanca, Costa Rica. Tesis de Mestría,
Centro Agronómico Tropical de Investigación y Enseñanza, Turrialba, Costa Rica,
125pp.
Goulart, F.V.B., N.C. Caceres, M.E. Graipel, M.A. Tortato, I.R. Ghizoni
Junior & L.G.R. Oliveira-Santos (2009). Habitat selection by large mammals in a southern Brazilian Atlantic
Forest. Mammalian Biology 74(3): 182–190; http://dx.doi.org/10.1016/j.mambio.2009.02.006
Head, J.S., M.M. Robbins, R. Mundry, L. Makaga & C. Boesch (2012). Remote video-camera traps measure habitat use and
competitive exclusion among sympatric Chimpanzee, Gorilla and elephant in
Loango National Park, Gabon. Journal of Tropical Ecology 28(06):
571–583; http://dx.doi.org/10.1017/S0266467412000612
Heithaus, M.R. & L.M. Dill (2002). Food availability and tiger shark predation risk influence Bottlenose
Dolphin Habitat Use. Ecology 83(2): 480–491; http://dx.doi.org/10.1890/0012-9658(2002)083[0480:FAATSP]2.0.CO;2
Henry, O. (1999). Frugivory
and the importance of seeds in the diet of the Orange-rumped Agouti (Dasyprocta
leporina) in French Guiana. Journal of Tropical Ecology 15(3):
291–300.
Hines, J. (2006). Presence
2.4 - Software to estimate patch occupancy and related parameters. USGS-PWRC.
Horn, H.S. (1966). Measurement
of overlap in comparative ecological studies. American Naturalist100(914): 419–424.
Isasi-Catalá, E. (2009).
Evaluación del estado de conservación del Jaguar (Panthera onca) y sus
principales presas naturales en el Parque Nacional Guatopo, estado Miranda, pp.
112. In: Giraldo D., F. Rojas-Suares & V. Romero (eds.). Una Mano a la
naturaleza, Conservando las especies amenazadas venezolanas. Provita y
Shell Venezuela S.A, Caracas, Venezuela.
Isasi-Catalá, E. (2012).
Estudio del estado de conservación del Jaguar (Panthera onca) en el
Parque Nacional Guatopo. Tesis Doctoral, Universidad Simón Bolívar, Sartenejas,
Venezuela, 393pp.
Isasi-Catalá, E. (2013).
Estado de conservación del Jaguar (Panthera onca) en el Parque Nacional
Guatopo, Venezuela: unidad prioritaria para su conservación, pp. 95–102. In:
Garrido E.P. & C. Castaño-Uribe (eds.). Grandes Felinos de Colombia,
Vol. I. Panthera Colombia, Fundación Herencia Ambiental Caribe,Conservación
Internacional y Cat Specialist Group UICN/SSC.
Johnson, D.H. (1980). The
comparison of usage and availability measurements for evaluating resource
preference. Ecology 61(1): 65–71; http://dx.doi.org/10.2307/1937156
Karanth, K. & S. Kumar (2002). Field surveys: assessing relative abundances of tigers and prey, pp.
71–85. In: Karanth K. & J. Nichols (eds.). Monitoring Tigers and Their
Prey: A Manual for Researchers, Managers and Conservationists in Tropical Asia.
Centre for Wildlife Studies, Bangalore, India.
Kays, R.W. & K.M. Slauson (2008). Remote cameras, pp. 110–140. In: Long R., P. MacKay, W. Zielinski
& J. Ray (eds.). Noninvasive Survey Methods for Carnivores. Island
Press, Washington DC, US.
Kuroiwa, A. & C. Ascorra (2002). Dieta y densidad de posibles presas de Jaguar en las inmediaciones de
la zona de reserva Tambopata-Candamo, Perú, pp. 199–208. In Medellin R., C.
Equihua, C. Chetkiewicz, P. Crawshaw, A. Rabinowitz, K.F. Redford, J. Robinson,
E. Sanderson & A. Taber (eds.). El Jaguar en El Nuevo Milenio. Fondo
de cultura económica, Universidad Nacional Autónoma de México, Wildlife
Conservation Society, México.
Linares, O. (1998). Mamíferos
de Venezuela. Sociedad Conservacionista Audubon de Venezuela, Caracas,
Venezuela, 610pp.
Linkie, M. (2008). Single-Species,
Single-Season: Occupancy Tutorials. DICE, University of Kent, UK, 25pp.
Linkie, M., Y. Dinata, A. Nugroho & I.A. Haidir (2007). Estimating occupancy of a data deficient mammalian
species living in tropical rainforests: Sun Bears in the Kerinci Seblat region,
Sumatra. Biological Conservation 137(1): 20–27; http://dx.doi.org/10.1016/j.biocon.2007.01.016
MacKenzie, D.I. & J.D. Nichols (2004). Occupancy as a surrogate for abundance estimation. Animal
Biodiversity and Conservation 27(1): 461–467.
MacKenzie, D.I., J.D. Nichols, G.B. Lachman, S. Droege, J.A. Royle &
C.A. Langtimm (2002). Estimating
site occupancy rates when detection probabilities are less than one. Ecology83(8): 2248–2255; http://dx.doi.org/10.1890/0012-9658(2002)083[2248:ESORWD]2.0.CO;2
MacKenzie, D.I., J.D. Nichols, J.A. Royle, K.H. Pollock, L.L. Bailey
& J.E. Hines (2006). Occupancy
Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence.
Academic Press, London, UK, 344pp.
Maffei, L., E. Cuéllar & A. Noss (2002). Uso de Trampas-Cámara para la Evaluación de Mamíferos en el Ecotono
Chaco-Chiquitanía. Revista
Boliviana de Ecología 11: 55–65.
Maffei,
L., A.J. Noss, E. Cuéllar & D.I. Rumiz (2005). Ocelot (Felis pardalis)
population densities, activity, and ranging behaviour in the dry forests of
eastern Bolivia: data from camera trapping. Journal of Tropical Ecology21(3): 349–353; http://dx.doi.org/10.1017/S0266467405002397
MARNR (1992). Área
naturales protegidas de Venezuela - Serie Aspectos Conceptuales y
Metodológicos. DGPOA/ACM/01.
Michalski, F. & D. Norris (2011). Activity pattern of Cuniculus paca (Rodentia: Cuniculidae) in
relation to lunar illumination and other abiotic variables in the southern
Brazilian Amazon. Zoologia 28(6): 701-708; http://dx.doi.org/10.1590/S1984-46702011000600002
Mostacedo, B. & T. Fredericksen (2000). Manual de Métodos Básicos de Muestreo y Análisis en Ecología
Vegetal. Editora El País, Santa Cruz, Bolivia, 87pp.
Negroes, N., R. Sollmann, C. Fonseca, A.T.A. Jácomo, E. Revilla & L.
Silveira (2012). One or two
cameras per station? Monitoring Jaguars and other mammals in the Amazon. Ecological
Research 27: 639–648; http://dx.doi.org/10.1007/s11284-012-0938-4
Noss, A., M. Kelly, H. Camblos & D. Rumiz (2006). Pumas y Jaguares simpátricos: Datos de
trampas-cámara en Bolivia y Belize. MEMORIAS: Manejo de Fauna silvestre en
Amazonia y Latinoamérica 229-237pp.
Nowak, R.M. (1991). Walker’s
Mammals of the World. 5th edition. The John Hopkins University Press,
Baltimore, US, 1615pp.
Perera-Romero, L. (2006).
Aporte para la estructuración de un Sistema de Información Geográfica del
Parque Nacional Guatopo. Informe Final de Cursos en Cooperación, Universidad
Simón Bolívar, Caracas, Venezuela.
Perez, E.M. (1992). Agouti
Paca. Mammalian Species Available online 11 December 2012;
http://www.science.smith.edu/departments/Biology/VHAYSSEN/msi/
Polisar,
J., I. Maxit, D. Scognamillo, L. Farrell, M.E. Sunquist & J.F. Eisenberg (2003). Jaguars,
Pumas, their prey base, and cattle ranching: ecological interpretations of a
management problem. Biological
Conservation 109(2): 297–310; http://dx.doi.org/10.1016/S0006-3207(02)00157-X
Queirolo, D., E. Vieira, L. Emmons & R. Samudio (2008). Cuniculus
paca. IUCN Red List of Threatened
Species Version 2012.2. Available online 26 March 2013; www.iucnredlist.org
Ríos-Uzeda, B., H. Gómez & R. Wallace (2007). A preliminary density estimate for Andean bear using
camera-trapping methods. Ursus 18(1): 124–128; http://dx.doi.org/10.2192/1537-6176(2007)18[124:APDEFA]2.0.CO;2
Rodríguez, J.P. & F. Rojas-Suárez (2008). Libro Rojo de la fauna Venezolana. PROVITA, S.A., Caracas,
Venezuela, 364pp.
Rovero, F., F. Zimmermann, D. Berzi & P. Meek (2013). Which camera trap type and how many do I need? A
review of camera features and study designs for a range of wildlife research
applications. Hystrix, the Italian Journal of Mammalogy 24(2): 148–156; http://dx.doi.org/10.4404/hystrix-24.2-8789
Royle, J.A. & J.D. Nichols (2003). Estimating abundance from repeated presence-absence data or point
counts. Ecology 84(3): 777–790; http://dx.doi.org/10.1890/0012-9658(2003)084[0777:EAFRPA]2.0.CO;2
Schipper, J. (2007). Camera-trap
avoidance by Kinkajous Potos flavus: rethinking the “non-invasive”
paradigm. Small Carnivore Conservation 36: 38–41.
Silva, J. & S. Strahl (1994). Usos folclóricos de la fauna silvestre en nueve parques nacionales al
norte de Venezuela. Vida Silvestre Neotropical 3(2): 100–107.
Silva, J. & S. Strahl (1996). La caza furtiva en los parques nacionales al norte de Venezuela. Vida
Silvestre Neotropical 5(2): 126–139.
Srbek-Araujo, A.C. & A. Garcia Chiarello (2005). Is camera-trapping an efficient method for surveying
mammals in Neotropical Forests? A case study in south-eastern Brazil. Journal
of Tropical Ecology 21: 121–125; http://dx.doi.org/10.1017/S0266467404001956
Sundell,
J., D. Dudek, I. Klemme, E. Koivisto, J. Pusenius & H. Ylonen (2004). Variation in
predation risk and vole feeding behaviour: a field test of the risk allocation
hypothesis. Oecologia 139(1): 157–162; http://dx.doi.org/10.1007/s00442-004-1490-x
Tarnaud, L. (2006).
Cathemerality in the Mayotte Brown Lemur (Eulemur fulvus): Seasonality
and food quality. Folia Primatologica 77(1-2): 166–177.
Tobler, M.W., S.E. Carrillo-Percastegui, R. Leite Pitman, R. Mares &
G. Powell (2008). An
evaluation of camera traps for inventorying large- and medium-sized terrestrial
rainforest mammals. Animal Conservation 11(3): 169–178; http://dx.doi.org/10.1111/j.1469-1795.2008.00169.x
Van Schaik, C.P. & M. Griffiths (1996). Activity periods of Indonesian rain forest mammals. Biotropica28: 105–112.
Venezuela (1958). Decreto de
creación del Parque Nacional Guatopo. Decreto múmero 122 (31 de Marzo de 1958),
Gaceta Oficial número 25624, INPARQUES.
Venezuela (1996a). Animales
Vedados para la Caza. Decreto 1485.
Venezuela (1996b). Especies en
Peligro de Extinción. Decreto 1486.
Wasserberg, G., B.P. Kotler & Z. Abramsky (2006). The role of site, habitat, seasonality and competition
in determining the nightly activity patterns of psammophilic gerbils in a
centrifugally organized community. Oikos 112(3): 573–579; http://dx.doi.org/10.1111/j.0030-1299.2006.14302.x
Weckel, M., W. Giuliano & S. Silver (2006). Cockscomb Revisited: Jaguar Diet in the Cockscomb
Basin Wildlife Sanctuary, Belize. Biotropica 38(5): 687–690; http://dx.doi.org/10.1111/j.1744-7429.2006.00190.x
Yerena, E. (1985).
Caracterización y Análisis de los Recursos Naturales del Parque Nacional
Guatopo: Zonificación y Bases para el Plan de Manejo Biológico. Trabajo
Especial de Grado, Universidad Simón Bolívar, Caracas, Venezuela, 600pp.